



Анаэробные организмы. Дрожжи аэробы или анаэробы
аэробы
аэробыАЭРОБНЫЕ ОРГАНИЗМЫ – организмы, нуждающиеся в молекулярном кислороде. Облигатные А. получают энергию только за счет аэробного дыхания, при котором кислород играет роль терминального окислителя. Облигатные А., нуждающиеся в пониженной концентрации кислорода в среде (порядка 2 %), получили название микроаэрофилы. Факультативные А. способны существовать как в кислородных, так и в бескислородных условиях, переключаясь с аэробного дыхания на брожение или анаэробное дыхание (дрожжи, энтеробактерии).
(Источник: «Микробиология: словарь терминов», Фирсов Н.Н., М: Дрофа, 2006 г.)
Аэробыоблигатные аэробные организмы, разнородная группа микроорганизмов, использующих аэробный (окислительный) тип биол. окисления субстратов. Метаболизм А. осуществляется только при наличии в среде обитания достаточно высокой концентрации свободного кислорода, к-рый выполняет функцию конечного акцептора отнятых от субстрата электронов. Окисление субстрата А. происходит в циклических процессах (цикле трикарбоновых к-т и его вариантах, пентозо-фосфатном цикле, цикле дикарбоновых к-т) с образованием воды и углекислого газа и высвобождением почти всей энергии, заключенной в окисляемом субстрате. Высвобожденная из субстрата энергия аккумулируется А. в макроэргических пирофосфатных связях: АТФ, НАДФ и др. У А. выявляется большинство ферментов дыхательной цепи, но для их дифференциации от анаэробов в основном используют определение оксидаз. Широко распространены в природе. Они составляют значительную долю нормальной микрофлоры животных, включая человека, и возбудителей инфекц. заболеваний. К ним, в частности, относятся вибрионы, микобактерии, псевдомонады, бациллы, фотобактерии, нитрифицирующие бактерии. Культивирование А. осуществляют на средах с полным доступом кислорода воздуха, однако наибольший выход биомассы можно получить при создании оптимальной концентрации кислорода, т. к. многие виды имеют характерные для них константы минимальной, максимальной и оптимальной концентрации кислорода.
(Источник: «Словарь терминов микробиологии»)
Поделитесь на страничкеslovar.wikireading.ru
Анаэробы строгие - Справочник химика 21
Микроорганизмы, которые погибают в присутствии кислорода, называют облигатными (строгими) анаэробами. Те микроорганизмы, которые могут существовать в аэробных и анаэробных условиях, называют факультативными (условными) анаэробами. Последние могут изменять тип дыхания в зависимости от среды (дрожжи). Анаэробные дыхательные процессы называют брожением. Это явление используется человеком для получения с помощью микроорганизмов ряда ценных продуктов этилового и бутилового спиртов, масляной, молочной и уксусной кислот и т. п. [c.16] Многие из анаэробных бактерий являются строгими анаэробами , т. е. вообще не переносят присутствия кислорода, как, например метаногенные бактерии . [c.145]Образование молекулярного кислорода из воды в процессе фотосинтеза явилось, несомненно, важнейшим событием в эволюции и имело далеко идущие последствия. По мере накопления кислорода в атмосфере Земли облигатные анаэробы (для которых кислород токсичен) оста-.лись только в строго анаэробных средах, уступив место новым классам -бактерий, обладающих механизмами детоксикации кислорода и использования его для окисления сложных органических соединений с целью получения необходимой энергии. [c.26]
Маслянокислые бактерии — строгие анаэробы, имеющие подвижные крупные спорообразующие палочки длиной до 10 мкм. Споры их цилиндрической илп эллипсоидальной формы. Наряду с масляной кислотой они могут образовывать (в меньших количествах) уксусную, молочную, капроновую, каприловую и друг е кислоты, а также этиловый и бутиловый спирты. Возбудители этого брожения развиваются главным образом в трубопроводах, насосах и других скрытых местах. Оптимальная температура для роста бактерий 30—40°С, при pH ниже 4,9 они ие развиваются. [c.210]
Метанобразующие бактерии — строгие анаэробы. Первые исследования чистых культур, вьщеленных из рубца жвачных животных, показали, что рост их возможен при начальном окислительно-восстановительном потенциале среды ниже -300 мВ. Рост некоторых видов полностью подавляется при содержании в газовой фазе более 0,004 % молекулярного кислорода. В последнее время, однако, описаны виды с относительно низкой чувствительностью к О2. В их клетках найдена супероксиддисмутаза. Возможно, в природе такие виды могут сохранять жизнеспособность при кратковременных контактах с О2 и возобновлять рост в анаэробных условиях. [c.424]
На следующей стадии эволюции появились, видимо, организмы, родственные современным фотосинтезирующим бактериям (пурпурным и зеленым) они могли использовать энергию солнечного света. Любопытно, что большинство этих (грамотрицательных) фотосинтезирующих бактерий—строгие анаэробы. В отличие от высших растений ни один из указанных микроорганизмов не выделяет кислорода. Напротив, для. восстановления двуокиси углерода в процессе фотосинтеза им необхо ДИМ водород, который они получают либо путем расщепления неорганических соединений типа h3S, тиосульфата или Нг, либо из органичен ских веществ. [c.25]
Группа 14. Грамположительные, не образующие спор палочки правильной формы. Группа — конгломерат, состоящий из 7 родов, объединенных несколькими общими морфологическими и физиологическими признаками клетки палочковидной формы (от кокковидных до удлиненных, одиночных или образующих цепочки), мезофилы строгие или факультативные аэробы, есть микроаэрофилы и аэротолерантные анаэробы хемоорганогетеротрофы, растущие только на сложных средах. [c.173]
Между ними существует переходная группа, которая получила название микроаэрофильных организмов. Для их жизнедеятельности требуются малые концентрации кислорода в среде. Анаэробные микроорганизмы подразделяются на а) факультативные (условные) анаэробы, которые могут развиваться как в присутствии, так и в отсутствии молекулярного кислорода и б) облигатные (безусловные, строгие) анаэробы, развивающиеся только в отсутствии молекулярного кислорода, который для них является ядом. Указанное деление микроорганизмов носит, конечно, условный характер. В зависимости от потребности в кислороде у микробов может быть два типа дыхания аэробное, или настоящее дыхание, и анаэробное дыхание, или брожение. Тип дыхания зависит также от наличия тех или иных дыхательных ферментов в микробной клетке. Остановимся более подробно на этих типах дыхания. [c.528]
В то же время для ряда цианобактерий показана способность расти на свету в строго анаэробных условиях. Это относится к видам, осуществляющим фотосинтез бескислородного типа, которых в соответствии с принятой классификацией следует отнести к факультативным анаэробам. (Фотосинтез любого типа по своей природе — анаэробный процесс. Это хорошо видно в случае фотосинтеза бескислородного типа и менее очевидно для кислородно- [c.316]
В отношении к молекулярному кислороду среди фототрофных эубактерий на одном полюсе располагаются строгие анаэробы, на другом — организмы, у которых О2 образуется внутриклеточно. Многие виды — факультативные анаэробы, есть аэротолерантные формы и микроаэрофилы. У фотосинтезирующих эубактерий молекулярный кислород часто выступает как могучий фактор, регулирующий их метаболизм в аэробных условиях у пурпурных и зеленых бактерий репрессируется синтез фотосинтетических пигментов и тем самым уничтожается основа для фототрофного способа существования. [c.324]
Обнаружение супероксиддисмутазы у строгих анаэробов (гораздо более распространенное, чем предполагали раньше) ставит вопрос о ее физиологической роли у этих организмов. Способность последних расти только в бескислородной среде делает неясным функции фермента в данных условиях. Возможно, что только при попадании строгого анаэроба в неблагоприятные для него аэробные условия синтез фермента индуцируется молекулярным кислородом, что и обеспечивает организму защиту от О2 в этих условиях. [c.335]
Тот факт, что все существующие на Земле прокариоты, даже строгие анаэробы, в присутствии О2 его поглощают, указывает на осуществление ими каких-то реакций взаимодействия с молекулярным кислородом. По отношению к О2 все прокариоты могут быть разделены на несколько физиологических групп (см. рис. 34). Такое подразделение говорит о необходимости или вреде молекулярного кислорода, но не раскрывает механизмов взаимодействия с ним клетки. Действительно, сейчас мы знаем, что О2 может быть необходим клетке для получения энергии или же для осуществления всего одной реакции, не имеющей энергетического значения. [c.344]
Для архебактерий как группы в целом характерна способность существовать в щироком диапазоне условий внещней среды. Среди них есть строгие и факультативные анаэробы и облигатные [c.415]
Группа 15. Грамположительные, не образующие спор палочки неправильной формы. Группа разнообразна. Большинство — фампо-ложительные палочки неправильной формы, растущие в присутствии воздуха и не образующие эндоспор, но есть в фуппе бактерии, имеющие форму кокков или палочек правильной формы, окрашивающиеся отрицательно по Граму и являющиеся строгими анаэробами. [c.173]
Возбудители маслянокислого брожения — строгие анаэробы, подвижные палочки с клОстридиальным или плектридиальным типом спорообразования. [c.97]
Поли-р-гидроксимасляную кислоту образуют многие аэробные бактерии (рис. 1.12), а также цианобактерии и анаэробные фототрофные бактерии. У факультативных анаэробов и строгих аэробов она накапливается, когда клетки испытывают недостаток О2 и переходят на брожение поэтому ее можно рассматривать как полимерный внутриклеточный продукт брожения. В аэробных условиях этот продукт может быть снова вовлечен в метаболизм как источник энергии и углерода и использован в процессе дыхания. [c.34]
Различают облигатных, т. е. строгих, безусловных анаэробов, для которых кислород вреден примером могут служить маслянокислые бактерии. Факультативные анаэробы (или условные) могут жить и в присутствии кислорода и без него. К факультативным анаэробам относятся молочнокислые бактерии, дрожжи. [c.129]
После работ Омелянского проводились систематические исследования механизма образования метана из органических и неорганических веществ. Сложность изучения метанообразующих микроорганизмов связана с тем, что оии являются строгими анаэробами, поэтому их чрезвычайно трудно изолировать. Кроме того, метановые бактерии очень медленно развиваются в культурах. Ряд исследователей связывают медленное развитие метановых бактерий в питательной среде с ее окислительно-восстановительными условиями. Установлена прямая зависимость механизма преобразования органического вещества от гНз среды. Так, при значении Ж2=12—12,9 разложение кальциевой соли муравьиной кислоты протекает с образованием водорода по следующей схеме (НС00)2Са-1-Н20->СаС0з + С02 + 2Н2. А при введении в систему газообразного водорода и значения гНг = 6—7 муравьиная кислота минерализуется с образованием метана по уравнению НСООН-Ь + ЗН2 >СН4 - - 2Н2О. [c.314]
У клостридий и у других строгих анаэробов обнаружен фермент, сходный с липоилзависимыми дегидрогеназами кетокислот — пируват ферредоксин — оксидоредуктаза, катализирующая обратимое декарбок- [c.272]
При осуществлении текущего санитарного надзора рекомендуется сокращенный анализ. Он включает определение ОМЧ, БГКП, титра строгих анаэробов, термофильных бактерий, нитрифицирующих бактерий. В полный санитарно-микробиологический анализ дополнительно входят определение численности и процентного отношения спор к общему количеству микроорганизмов, количество актиномицетов, грибов, целлюлозоразрушающих микроорганизмов и аммонификаторов, основных групп почвенного микробиоценоза и ряд дополнительных исследований (например, токсичность почв для микроорганизмов). По эпидемическим показаниям в ходе исследований проводят обнаружение патогенных микроорганизмов (табл. 11.2). [c.421]
Объединены прокариоты, по имеющимся данным, претендующие на более раннее происхождение, чем формы, включенные в I и П отделы. Клетки разной формы кокки, палочки, нити. Многие плеоморфны. Большинство имеют клеточную стенку, но она не содержит типичного пептидогликана. Клеточная стенка может быть построена только из белковых макромолекул или гетерополисахаридов. Окрашивание по Граму отрицательное или положительное. Большинство — строгие анаэробы. Многие имеют жгутики. Характеризуются экологическим и метаболическим разнообразием, способностью жить в экстремальных условиях. Объединены в класс АгсЬаеоЬас1ег1а [c.159]
Группа 7. Бактерии, характеризующиеся диссимиляционным восстановлением серы или сульфата. В составе группы эубактерии с разной морфологией и следующими одинаковыми свойствами грамотрицательные строгие анаэробы, использующие в качестве акцептора электронов молекулярную серу или ее окисленные соединения, которые восстанавливаются при этом до HjS. Некоторые виды способны к брожению. Есть среди них азотфиксаторы. [c.168]
К V подгруппе отнесены архебактерии, характеризующиеся совокупностью следующих признаков облигатные термофилы ацидофилы или нейтрофилы аэробы, факультативные или строгие анаэробы автотрофы или гетеротрофы. Метаболизм больщинства из них связан с молекулярной серой (S ). Представители порядка Thermoproteales в процессе хемолитоавтотрофного роста получают энергию в реакции [c.180]
Большинство существующих облигатных анаэробов среди эубактерий имеют вторичное происхождение как следствие повторной адаптации к анаэробным условиям, сопровождающейся, как правило, изменениями дефадационного характера потерей способности взаимодействовать с О2, утратой некоторых компонентов переноса электронов, большей зависимостью от готовых органических соединений среды обитания и т.д. Примером могут служить строго анаэробные эубактерии, составляющие основную микрофлору рубца и пищеварительного тракта животных и человека. Это в большинстве фамотрицательные кокки или палочки, способные сбраживать сахара и/или аминокислоты. У многих из них обнаружены цитохромы Ьиаи показана способность синтезировать АТФ по механизму мембранзависимого фосфорилирования. [c.261]
В течение длительного времени пурпурные серобактерии считали строгими анаэробами и облигатными фототрофами. Недавно было показано, что спектр отношения к молекулярному кислороду в этой группе достаточно широк. В большинстве пурпурные серобактерии высоко чувствительны к О2, однако и среди них есть виды, растущие в темноте в аэробных условиях на минеральной среде или с использованием органических соединений. Хемолито-автотрофный рост при низком содержании О2 обнаружен у ряда пурпурных серобактерий, ассимилирующих СО2 в восстановительном пентозофосфатном цикле, а энергию получающих в процессе дыхания в результате окисления сульфида, тиосульфата, молекулярной серы или Н2. Для некоторых представителей родов E tothiorhodospira и Thio apsa показан аэробный хемоорганогетеротрофный рост. Органические вещества в этом случае используются как источники углерода и энергии. Последняя запасается в процессе дыхания. Некоторые пурпурные серобактерии оказались также способными расти в темноте в анаэробных условиях (в атмосфере аргона или молекулярного водорода), сбраживая некоторые сахара или органические кислоты. [c.299]
В течение длительного времени зеленые бактерии принимали за зеленые или сине-зеленые водоросли (цианобактерии). Начало их изучения как бактерий связано с именами С. Н. Виноградского и К. ван Ниля. Эта небольшая группа эубактерий, осушествляющих фотосинтез бескислородного типа, разделена на две подгруппы. Зеленые серобактерии — строгие анаэробы и облигатные фототрофы, способные расти на среде с Н28 или молекулярной серой в качестве единственного донора электронов при окислении сульфида до молекулярной серы последняя всегда откладывается вне клетки. [c.302]
Все зеленые серобактерии — облигатные фотолитоавтотрофы и строгие анаэробы (гораздо более строгие, чем пурпурные серобактерии). В присутствии О2 они не растут. Основной источник углерода — углекислота. Как доноры электронов могут использовать [c.303]
Среди облигатных анаэробов супероксиддисмутаза обнаружена у многих представителей рода lostridium. Изучение их устойчивости к О2 обнаруживает четкую связь с содержанием в клетках этого фермента. Виды, имеющие супероксиддисмутазу, характеризуются умеренной или даже высокой устойчивостью к О2 по сравнению с видами, у которых этот фермент отсутствует. Супер-оксидцисмутаза найдена у разных видов строго анаэробных бактерий. Число организмов с не выявленной до сих пор супероксид-дисмутазой очень мало. [c.335]
Каталазная и пероксидазная активности обнаружены у всех облигатно и факультативно аэробных прокариот. Среди облигатных анаэробов эти ферменты распространены значительно в меньщей степени, чем супероксиддисмутаза. Обнаружены многие строгие и аэротолерантные анаэробы, содержащие супероксиддисму- [c.336]
Все сульфатвосстанавливающие бактерии — облигатные анаэробы. Многие из них относятся к категории строгих анаэробов, для роста которых требуется не только отсутствие Oj, но и низкий окислительно-восстановительный потенциал среды. В то же время некоторые штаммы проявляют устойчивость к Оз и вьгживают при разной длительности аэрирования среды. [c.389]
Таким образом, отношение к О2 в группе серозависимых архебактерий охватывает диапазон от облигатного аэробиоза до строгого анаэробиоза. Есть и факультативные формы. Облигатно аэробные формы осуществляют окисление 8°, строгие анаэробы — только восстановление, факультативные — в зависимости от условий могут окислять или восстанавливать 8 . [c.435]
Почти все микроорганизмы, окисляющие С -содинения, не нуждаются в витаминах и других факторах роста. Известны также строгие анаэробы, растущие в автотрофных условиях, окисляя молекулярный водород. [c.153]
Смешанная культура метанообразующих бактерий. Эту культуру термофильных метанообразующих бактерий используют для получения витаминизированных кормовых добавок для сельскохозяйственных животных. Эта технология принципиально отличается от той, которая была описана для пропионовокислых бактерий. Метанообразующие бактерии - строгие анаэробы, отличаются большой чувствительностью к кислороду, плохо переносят перемешивание и с большим трудом выделяются в чистую культуру. Бактерии Methanoba terium kuznetzeovii используют метанол с образованием ацетата в качестве промежуточного продукта. [c.293]
По типу дыхания все бактерии можно разделить на три группы 1) строгие анаэробы — маслянокислые бактерии, возбудители ботулизма 2) строгие аэробы — уксуснокислые бактерии, палочки инфлуэнцы, псевдомонасы 3) факультативные анаэробы — кишечная палочка, молочнокислые бактерии, по-давляюшее большинство патогенных бактерий и дрожжи. У представителей всех этих трех групп возможно дыхание путем непрямого окисления — дегидрогенирования. С отщеплением водорода освобождается электрон, а энергия поступает на нужды микробной клетки. Например, некоторые водные микроорганизмы осуществляют реакцию [c.94]
Первая стадия заключается в разложении крупных органических соединений и превращении их в органические кислоты с выделением газообразных побочных продуктов углекислого газа, метана и следов сероводорода. Эта стадия осуществляется разнообразными факультативными бактериями, функционирующими в лишенной кислорода среде. Если бы процесс на этом остановился, скопившиеся кислоты привели бы к понижению pH и к ингибированию последующего распада при закисании оставшихся сырых сточных вод. Для того чтобьг произошло сбраживание, на второй стадии необходима газификация для превращения органических кислот в метан и углекислый газ. Бактерии, расщепляющие кислоты с образованием метана, являются строгими анаэробами и очень чувствительны к условиям окружающей среды, т. е. к температуре, pH и анаэробиозу. [c.340]
По отношению к кислороду в среде, где происходит вегетация, микроорганизмы делятся на облигатные (строго обязательные) аэробы, облигатные (строго обязательные) анаэробы, факультативные анаэробы (растущие в аэробных и в анаэробных условиях), микроаэрофилъные анаэробы (переноснш,ие малое количество кислорода). [c.22]
chem21.info
Брожение анаэробное среды - Справочник химика 21
Выше мы разобрали наиболее простой способ решения донор-акцепторной проблемы, который реализуется в виде молочнокислого брожения у группы гомоферментативных молочнокислых бактерий. Дальнейшие поиски на путях эволюции привели к формированию других метаболических возможностей для решения этой проблемы. Одна из них заключается в том, что из пировиноградной кислоты в результате ее окислительного декарбоксилирования образуется ацетальдегид, который становится конечным акцептором водорода. В итоге из 1 молекулы гексозы образуются 2 молекулы этилового спирта и 2 молекулы углекислоты. Процесс получил название спиртового брожения. Спиртовое брожение распространено среди прокариотных (различные облигатно и факультативно анаэробные эубактерии) и эукариотных (дрожжи) форм. В анаэробных условиях у высших растений также отмечено накопление этилового спирта. [c.219] А. Обычная обработка. Наиболее важную, если не основную, роль в обработке стоков играют микробиологические процессы, основанные на усиленной минерализации органического вещества (например, обработка активированным илом, капельные фильтры, медленные песчаные фильтры) и на метановом брожении (анаэробное разложение). Основные конечные продукты этих процессов или безопасны (двуокись углерода, вода), или легко удаляются (клетки, СН4). Так как клетки патогенных микроорганизмов в чужеродной для них среде также представляют органическое вещество, подлежащее минерализации, биологическая обработка ведет к снижению их количества. [c.289]Обычно рекомендуется, чтобы брожение велось в закрытых танках из нержавеющей стали, что позволяет обеспечить анаэробную среду, позволяющую снизить потери летучих соединений. [c.444]
Микроорганизмы, которые погибают в присутствии кислорода, называют облигатными (строгими) анаэробами. Те микроорганизмы, которые могут существовать в аэробных и анаэробных условиях, называют факультативными (условными) анаэробами. Последние могут изменять тип дыхания в зависимости от среды (дрожжи). Анаэробные дыхательные процессы называют брожением. Это явление используется человеком для получения с помощью микроорганизмов ряда ценных продуктов этилового и бутилового спиртов, масляной, молочной и уксусной кислот и т. п. [c.16]
Пропионовую кислоту получают в анаэробных условиях методом глубинного культивирования (рис. 51). Используют среду, содержащую 2% глюкозы и источник органического азота, как, например, дрожжевой экстракт, а также соли молочной кислоты. Процесс идет в нейтральной среде (pH 6,8—7,2), при температуре 30°С, длится 7—12 сут. В процессе брожения накапливается про-пионовая, уксусная кислоты (5 1) и выделяется углекислый газ. Примерно 75% сахара потребляется на образование кислот, а 20% — на образование углекислого газа. [c.147]
Влияние окружающей среды на анаэробное брожение [c.153]
Между ними существует переходная группа, которая получила название микроаэрофильных организмов. Для их жизнедеятельности требуются малые концентрации кислорода в среде. Анаэробные микроорганизмы подразделяются на а) факультативные (условные) анаэробы, которые могут развиваться как в присутствии, так и в отсутствии молекулярного кислорода и б) облигатные (безусловные, строгие) анаэробы, развивающиеся только в отсутствии молекулярного кислорода, который для них является ядом. Указанное деление микроорганизмов носит, конечно, условный характер. В зависимости от потребности в кислороде у микробов может быть два типа дыхания аэробное, или настоящее дыхание, и анаэробное дыхание, или брожение. Тип дыхания зависит также от наличия тех или иных дыхательных ферментов в микробной клетке. Остановимся более подробно на этих типах дыхания. [c.528]
Для лучшего размножения, а также активации ослабленных дрожжей необходима аэрация питательной среды, так как построение веществ протоплазмы новых молодых клеток связано с затратой энергии, получаемой дрожжами в процессе дыхания. Поэтому небольшое количество воздуха подают обычно в начале брожения. Процесс же спиртового брожения происходит в анаэробных условиях, и необходимую для жизнедеятельности энергию дрожжи получают в процессе брожения. [c.543]
Под пробкой не должны оставаться пузырьки воздуха. В трубку над средой наливают вазелиновое масло небольшим слоем, и таким образом в сосуде с жидкой средой создаются анаэробные условия. Отсутствие углеводов в среде исключает процесс брожения. В этих условиях будут развиваться лишь микроорганизмы, способные использовать кислород связанных соединений (в первую очередь нитрата). [c.121]
О2 (последний не участвует в осуществляемых ими метаболических реакциях), но способные расти в его присутствии, являются по типу осуществляемого ими метаболизма облигатными анаэробами, устойчивыми к О2 внешней среды. Примером таких организмов служат молочнокислые бактерии. Многие прокариоты, относящиеся к этой же группе, приспособились в зависимости от наличия или отсутствия О2 в среде переключаться с одного метаболического пути на другой, например с дыхания на брожение, и наоборот. Такие организмы получили название факультативных анаэробов, или факультативных аэробов. Представителями этой физиологической группы прокариот являются энтеробактерии. В аэробных условиях они получают энергию в процессе дыхания. В анаэробных условиях источником энергии для них служат процессы брожения или анаэробного дыхания. [c.129]
В природе есть много мест с полным или почти полным отсутствием молекулярного кислорода. Это глубокие слои воды, почвы, илы морей и континентальных водоемов. Особую экологическую нишу для развития анаэробов представляют рубец и кишечник животных и человека. Облигатно анаэробный способ существования широко распространен среди эубактерий. Систематическое изучение анаэробных эубактерий, предпринятое в последние десятилетия, обнаружило неоднородность входящих в эту фуппу организмов, способных получать энергию в процессах брожения, фотосинтеза и анаэробного дыхания. [c.261]
Рост в темноте на органических средах за счет аэробного дыхания анаэробного дыхания брожения — + + 7 [c.305]
Могут ли экстремальные галофилы расти за счет энергии света или он служит только дополнительным источником энергии В лаборатории был показан рост этих бактерий в анаэробных условиях, исключающих возможность осуществления брожения или анаэробного дыхания, при освещении клеток, если они содержат значительные количества бактериородопсина или если в среду внесен ретиналь, для синтеза которого необходим молекулярный [c.422]
У анаэробных микроорганизмов акцептором водорода могут служить молекулы органических веществ с ненасыщенными связями. Такой тип дыхания называется брожением. Среди конечных продуктов брожения всегда имеются не полностью окисленные вещества, и обычно типы брожений получают свое название по конечным продуктам [c.128]
В отличие от синтеза АТФ в митохондриях синтез АТФ в анаэробной фазе распада углеводов (например, при спиртовом брожении или анаэробном дыхании) может иметь место и в гомогенной среде, и он не связан с клеточными структурами. [c.174]
До тех пор, пока всеобъемлющий термин биотехнология не стал общепринятым, для обозначения наиболее тесно связанных с биологией разнообразных технологий использовали такие названия, как прикладная микробиология, прикладная биохимия, технология ферментов, биоинженерия, прикладная генетика и прикладная биология. Если не принимать в расчет производства мыла, то первая же из числа возникших технологий такого рода стала предшественницей прикладной микробиологии. Наши предки не имели представления о процессах, лежащих в основе таких технологий. Они действовали скорее интуитивно, но в течение тысячелетий успешно использовали метод микробиологической ферментации для сохранения пищи (например, при получении сыра или уксуса), улучшения вкуса (например, хлеба и соевого соуса) и производства спиртных напитков. Пивоварение до сих пор остается наиболее важной (в денежном исчислении) отраслью биотехнологии. Во всем мире ежегодно производится около 10 литров пива стоимостью порядка 100 млн, фунтов стерлингов. В основе всех этих производств лежат реакции обмена веществ, происходящие при росте и размножении некоторых микроорганизмов в анаэробных условиях. В конце XIX в. благодаря трудам Пастера были созданы реальные предпосылки для дальнейшего развития прикладной (технической) микробиологии, а также в значительной мере и биотехнологии. Пастер установил, что микробы играют ключевую роль в процессах брожения, и показал, что в образовании отдельных продуктов участвуют разные их виды. Его исследования послужили основой развития в конце XIX и начале XX вв. бродильного производства органических растворителей (ацетона, этанола, бутанола и изопропанола) и других химических веществ, где использовались разнообразные виды микроорганизмов. Во всех этих процессах микробы в бескислородной среде осуществляют превращение углеводов растений в ценные продукты. В качестве источника энергии для роста микробы в этих условиях используют изменения энтропии при превращениях веществ. Совсем иначе обстоит дело в аэробных процессах при контролируемом окислении химических веществ до углекислого [c.11]
Прокариоты морфологически относительно слабо дифференцированы, поэтому среди них можно различить лишь ограниченное число форм. В основном это либо сферические формы, либо прямые и изогнутые палочки. С таким внешним единообразием удивительно контрастирует чрезвычайное многообразие и пластичность метаболических процессов. В то время как животные и растения нуждаются в молекулярном кислороде, многие группы прокариот способны жить без доступа воздуха (в анаэробных условиях), получая необходимую для роста энергию в результате брожения или анаэробного дыхания. Другие группы прокариот обладают способностью использовать энергию света и строят нужные им вещества либо из органических соединений, либо из углекислоты (двуокиси углерода). Некоторые бактерии могут получать энергию путем окисления различных неорганических соединений или элементов. Среди бактерий широко распространена также способность к фиксации молекулярного азота. [c.12]
Чистоту культур сульфатвосстанавливающих бактерий проверяют согласно методике, предложенной Постгейтом [195]. Исследуемую культуру высевают в среду с МПА для обнаружения факультативно-анаэробных гетеротрофных спутников в среде с сахарами и пептоном - для бактерий, осуществляющих брожение на среду Бейеринка [48] - для выявления тионовых бактерий. Для достоверности получаемые данные всегда необходимо сравнивать с контролем (стерильная среда). [c.62]
Решающее значение для совершенствования энергетических механизмов клеток имело появление в ходе эволюции способности к активному трансмембранному переносу ионов Н . АТРазная -помпа (рис. 3.29,1) должна была функционировать уже у самых примитивных клеток — протобионтов — для удаления избытка ионов Н+, которые накапливались в них при брожении (анаэробном окислении глюкозы). В результате выкачивания ионов Н+ во внутриклеточной среде не только поддерживался оптимальный для синтетических процессов уровень pH, но и возникал электрохимический мембранный потенциал, который стал энергетической основой мембранного транспорта и осморегуляции (рис. 3.29, II). [c.122]
Превращения углеводов, входящих в состав отмерших организмов, начинаются еще в водной среде. В верхнем слое донного осадка, в аэробной обстановке, может быть полностью минерали- зована даже клетчатка — наиболе устойчивая среди полисахаридов. В анаэробных условиях, когда происходят различные виды брожений, выделяются Н2О, СО2, СН4 и Нг. Микроорганизмы, использующие в качестве питания углеводы, синтезируют другие [c.30]
В первом случае процесс прекращают, когда достигнуто максимальное количество биомассы, когда максимально использован углерой и азот среды. Если источник углерода для образования биомассы и какого-либо продукта общий, например спиртовое брожение на сахарозе или мальтозе, тогда на определенном этапе увеличение биомассы ограничивают (при спиртовом брожении — путем образования факультативно анаэробных условий). [c.66]
Продуцентами этих кислот могут быть бактерии, плесневые грибы или дрожжи. Микроорганизмы, продуцирующие молочную кислоту, а также вызывающие спиртовое брожение, в ходе эволюции приспособились к анаэробному образу жизни. Уксусная и лимонная кислоты в свою очередь образуются в аэробных условиях. По-видимому, кислоты играют определенную роль в борьбе с конкурирующей микрофлорой, а также являются резервными источниками углерода. Так, Aspergillus niger после использования сахара могут использовать в качестве субстрата лимонную кислоту. В свою очередь уксуснокислые бактерии при отсутствии спирта в среде ассимилируют уксусную кислоту, окисляя ее до воды и СО2. [c.143]
Бактериальная микрофлора (рис. 139) представлена.следующими микроорганизмами 1) уксуснокислые бактерии, превращающие этиловый спирт в уксусную кислоту 2) молочнокислые бактерии, относящиеся к бесспо овым палочкообразным видам оптимальная температура для их развития 24—50° они анаэробны, используют сахар, превращая его в молочную кислоту и ряд других веществ (уксусная кислота, этиловый спирт) в результате жизнедеятельности молочнокислых и уксуснокислых бактерий значительно повышается кислотность сусла и бражки 3) маслянокислые и другие спороносные бактерии, использующие сахар (встречаются реже), а также сардины. Сардины представляют собой клетки, состоящие из восьми шариков, очень аэробны, превращают сахар в молочную и уксусную кислоты. Их можно обнаружить в сусле и бражке, полученных в результате гидролиза сельскохозяйственных отходов. Особенно благоприятной средой для развития инфекции служат хлопковые гидролизаты, богатые азотистыми и минеральными веществами. Маслянокислые бактерии являются довольно опасными врагами брожения, так как образуемая ими масляная кислота действует угнетающим образом на дрожжи [c.557]
Основными продуцентами этилового спирта, имеющими широкое практическое применение, являются дрожжи — одноклеточные эукари-отные микроорганизмы, принадлежащие к разным классам высших грибов. Наиболее распространенный способ размножения дрожжей — почкование. Дрожжи — аэробы со сформированным аппаратом дыхания, но в анаэробных условиях осуществляют спиртовое брожение по пути, рассмотренному в предыдущем разделе, т.е. получают энергию за счет субстратного фосфорилирования. Конструктивный метаболизм дрожжей основан на их хорошо развитых биосинтетических способностях. Есть виды дрожжей, развивающиеся на простых синтетических средах эти дрожжи способны синтезировать все необходимые им сложные органические соединения. Существуют виды, нуждающиеся в определенных витаминах группы В. Добавление к питательной среде веществ, содержащих комплекс витаминов, аминокислот, сахаров, приводит, как правило, к заметному стимулированию роста дрожжей. [c.223]
Свойство предельной окисленности молекулы СО2 используется в энергетическом метаболизме ряда анаэробных эубактерий, получающих энергию в процессе брожения, где СО2 служит для удаления избытка восстановителя, т.е. как конечный акцептор электронов. Эта же особенность молекулы СО2 находит применение и в энергетическом метаболизме некоторых анаэробных эубактерий (ацетогены) и архебактерий (метаногены), но у них электроны на СО2 поступают через цепь связанных с мембраной переносчиков электронного транспорта. СО2, участвующая в реакциях энергетического метаболизма, не включается в вещества клетки, а продукты ее восстановления (в виде молекул формиата, ацетата, метана) накапливаются в среде. В наибольшей степени способность вовлекать СО в метаболизм среди первично анаэробных хемогете-ротрофных эубактерий проявляется в фуппе клостридиев. [c.291]
Процесс брожения можно осуществлять в различных вариантах — непрерывном, двухстадийном. На рис. 128 показан двухстадийный процесс, когда в дрожжегенераторах (позиция 6) среду аэрируют 3—4 mVm 4 при 28—30°С и pH 4,2—4,5 дрожжи наращивают до 2,5—6,5% на сукие вещества, затем передают в бродильные аппараты (позиции 8,9), где в анаэробных условиях протекает спиртовое брожение. Перегонку или дистилляцию спирта проводят с целью его отделения от примесей и последующей ректификации до получения 96% этанола или абсолютного (100%) спирта. [c.402]
При использовании метилотрофами многоуглеродных субстратов этот процесс сопряжен с действием дыхательных электронотранспортных систем. Способность к росту при анаэробных условиях (при брожении) при наличии органических веществ в среде у факультативных анаэробных метилотрофов не обнаружена. [c.179]
Биологическая обработка — самый эффективный способ удаления органических веществ из городских сточных вод. Действие биологических очистных систем основано на том, что смешанные культуры микробов разлагают и удаляют коллоидные и растворенные органические вещества из раствора. Параметры среды, в которой находятся микроорганизмы в очистном сооружении, постоянно контролируются например, активный ил в достаточном количестве снабжается кислородом для поддержания аэробных условий. Сточная вода содержит биологическую пищу, питательные вещества для роста и микроорганизмы. Лица, незнакомые с очисткой сточных вод, часто спрашивают, откуда получают специальные биологические культуры. Многочисленные разновидности бактерий и простейших, присутствующие в бытовых сточных водах, служат на очистных установках в качестве исходной биологической затравки. Затем посредством тщательного контроля расхода поступающих сточных вод, рециркуляции микроорганизмов после их осаждения, снабжения кислородом и применения других способов удается вывести желательные биологические культуры, которые сохраняются для обработки загрязненных стоков. Биопленку на поверхности загрузки биофильтра получают, пропуская сточную воду через фильтр. Через несколько недель фильтр может работать, удаляя органические вещества из сточной жидкости, орошающей фильтр. Активный ил в механической или диффузно-воздушной системе начинает действовать при включении аэраторов и подаче сточной воды. Первоначально необходима высокая степень рециркуляции отстоя со дна вторичного отстойника для сохранения в достаточном количестве биологической культуры. Однако через короткий промежуток времени созревает устойчивый активный ил, который эффективно извлекает органические вещества из сточной воды. При включении в работу анаэробного сооружения приходится преодолевать более существенные затруднения, так как метанообразующие бактерии, необходимые для протекания процесса брожения, немногочисленны в необработанной сточной воде. Кроме того, эти анаэробы растут очень медленно и требуют оптимальных условий окружающей среды. Пуск анаэробной установки может быть значительно ускорен при заполнении тенка сточной водой и засеве ее достаточным количеством бродящего ила из близлежащей очистной установки. Сырой осадок сначала подают с незначительной дозой загрузки, а для поддержания должного значения pH в метантенк в необходимых количествах вводят известь. Даже при этих условиях проходит несколько месяцев, прежде чем установка начинает работать на полную мощность. [c.84]
Температура сбраживания является одним из важнейших факторов, влияющих на скорость распада осадка в метан-тенках. Большинство метан-тенков эксплуатируется в условиях мезофильного брожения при температуре 30—35°. При этом распад органического вещества осуществляется теми же группами микроорганизмов, которые вызывают его в двухъярусном отстойнике, работающем обычно при температуре 10—20°. Однако повышение температуры не только ускоряет процесс распада, но делает его более глубоким. При термофильном брожении (50—55°) в процессе распада принимает участие специфическая группа анаэробных термофильных микроорганизмов, имеющих весьма энергичный обмен вследствие высокой активности присущих им ферментов. Среди анаэробов термофилов имеются аммонифицирующие бактерии, денитрифицирующие и десульфурирующие, сбраживающие углеводы, разлагающие клетчатку и жирные кислоты, т. е. все те физиологические группы микробов, которые в условиях мезофильного брожения участвуют в разложении осадка. [c.218]
Судьба дигидрокодегидразы изменяется в зависимости от того, происходит ли реакция в анаэробной или аэробной среде. В нервом случае KoIHj отдает водород акцептору (например, ацетальдегиду в процессе спиртового брожения), регенерируя Kol (и образуя этанол). [c.801]
Для того чтобы выявить последовательность химических реакций, составляющих тот или иной метаболический путь, можно воспользоваться тремя главными экспериментальными подходами. Первый из них, наиболее прямой, заключается в изучении метаболического пути in vitro (в пробирке), т. е. не в самой живой ткани, а в ее бесклеточном экстракте, сохраняющем способность катализировать весь исследуемый процесс в целом. Еще в середине прошлого века стало, например, известно, что дрожжи сбраживают глюкозу до этилового спирта и СО2. Однако изучение отдельных стадий этого метаболического пути, поставляющего анаэробным дрожжевым клеткам почти всю необходимую им энергию, началось по-настоящему только с 1898 г., когда Эдуард Бухнер обнаружил, что отжатый из дрожжей сок, не содержащий живых клеток, тоже способен сбраживать глюкозу до этилового спирта и СО2 (разд. 9.1). Позже выяснилось, что брожение в таких экстрактах происходит лишь при добавлении неорганического фосфата и что по мере потребления глюкозы этот фосфат исчезает из экстракта. Оказалось, что в среде накапливается при этом какое-то фосфорилиро-ванное производное гексозы, обладающее всеми теми свойствами, какими должен обладать один из промежуточных продуктов на пути превращения глюкозы в этиловый спирт и СО2. После того как этот промежуточный продукт был идентифицирован, в дрожжевом экстракте удалось обнаружить фермент, превращающий его в другой продукт. Этот последний в свою очередь был выделен и идентифипдрован. Таким образом, идентифицированными оказались уже два промежуточных продукта расщепления глюкозы. Добавляя к эстрак-там ингибиторы ферментов, исследователи добивались накопления других промежуточных продуктов. В конце концов благодаря комбинированию такого рода приемов удалось выделить и идентифи- [c.391]
О 2 для них токсичен см. в конце раздела 7.4). Факультативные анаэробы растут как в присутствии, так и в отсутствие Oj. Среди них следует различать два типа аэротолерантные молочнокисглые бактерии могут расти в присутствии атмосферного кислорода, но не способны его использовать-они получают энергию исключительно с помощью брожения другие факультативно-анаэробные бактерии (Enteroba teria eae) и многие дрожжи могут переключаться с дыхания (в присутствии О2) на брожение (в отсутствие О2). [c.178]
Некоторые микроорганизмы, образующие при брожении кислоты, объединяют в одну физиологическую группу на том основании, что характерным, хотя и не главным продуктом брожения является у них муравь иная кислота. Наряду с муравьиной кислотой такие бактерии вьщеляют и некоторые другие кислоты такой тип метаболизма называют поэтому муравьинокислым брожением или брожением смешанного типа. Так как некоторые типичные представители этой группы обитают в кишечнике, все семейство носит название Enteroba teria eae. Это грам-отрицательные, активно подвижные, не образующие спор палочки с перитрихальным жгутикованием. Будучи факультативными аэробами, они обладают гемопротеинами (цитохромами и каталазой) и способны получать энергию как в процессе дыхания (в аэробных условиях), так и в процессе брожения (в анаэробных условиях). В отнощении питания эти бактерии исключительно нетребовательны-растут на простых синтетических средах, содержащих минеральные соли, углеводы и аммоний. Сбраживание глюкозы у всех представителей этой группы происходит с образованием кислот. Значение Enteroba teria eae для эпидемиологии, а также для разного рода экспериментальных исследований общеизвестно поэтому полезно будет рассмотреть здесь некоторых представителей этого семейства. [c.283]
На примере голомиктического озера можно описать биологические процессы, которые приводят к летнему расслоению и продолжаются несколько месяцев. В пронизанном лучами света эпилимнионе фитопланктон (диатомеи, жгутиковые, зеленые водоросли, цианобактерии) продуцирует биомассу. Обычно из окружающей среды в озеро поступает дополнительный органический материал. Часть этого органического вещества, в особенности частицы, содержащие целлюлозу, опускается на дно озера и разлагается. В начальной аэробной стадии разложения расходуется кислород, и на дне создаются анаэробные условия. В результате анаэробного распада образуются органические продукты брожения Н2, НдЗ, СН4 и СО2. Поскольку конвекции не происходит, эти продукты поступают из донных отложений в толщу воды очень медленно. Один только метан-главный продукт анаэробной цепи питания в донных осадках - выделяется в виде пузырьков газа. На своем пути к поверхности водоема часть метана переходит в раствор и окисляется использующими этот газ аэробными бактериями. Быстрое потребление кислорода в гиполимнионе обусловлено ускоренным распределением метана и ростом метанокисляющих бактерий. В конце концов во всем гиполимнионе создаются анаэробные условия. [c.507]
chem21.info
Дыхание дрожжей. Микробиология и биохимия вина.
Дыхание дрожжей
Каждый организм для осуществления процессов, совокупность которых составляет обмен веществ, нуждается в постоянном притоке энергии. Источником этой энергии является процесс диссимиляции.
Одни организмы получают эту энергию в результате окисления неорганических веществ (хемосинтезирующие микроорганизмы), другие в результате окисления органических веществ (аэробное дыхание), третьи в результате анаэробного распада углеводов (брожение).
По типу дыхания микроорганизмы можно разделить на аэробы и анаэробы. Первые получают энергию в результате окисления различных органических веществ (углеводов, спиртов, органических кислот), вторые - в результате анаэробного распада органических веществ. Анаэробы делятся в свою очередь на факультативные и облигатные. Факультативные могут жить как в присутствии кислорода, так и без него, а облигатные только в отсутствие кислорода.
Количество энергии, освобождающейся при дыхании, зависит от используемого материала и степени его окисления. Наибольшее количество энергии (2820 кДж) освобождается при аэробном дыхании (плесневые грибы, пленчатые дрожжи, уксусно- и молочнокислые бактерии).
Дрожжи по типу дыхания относятся к факультативным анаэробам, т. е. они могут существовать как в присутствии кислорода, так и без него. Необходимую для жизнедеятельности энергию они добывают в процессе анаэробного разложения сахара (брожение):
С6Н12О6 → 2С2Н5ОН + 2СО2 + 113,13 кДж.
Присутствие кислорода подавляет спиртовое брожение и дрожжи получают энергию путем кислородного дыхания (хересные дрожжи в процессе хересования вина).
Микроорганизмы используют для своих нужд не более 10-25% энергии, освобождающейся при дыхании. Остальная энергия выделяется в окружающую среду в виде тепла. Именно поэтому температура бродящего сусла выше температуры окружающей среды.
vinobio.narod.ru
Анаэробные организмы - это... Что такое Анаэробные организмы?
Аэробные и анаэробные бактерии предварительно идентифицируются в жидкой питательной среде по градиенту концентрации O2:1. Облигатные аэробные (нуждающиеся в кислороде) бактерии в основном собираются в верхней части пробирки, чтобы поглощать максимальное количество кислорода. (Исключение: микобактерии — рост пленкой на поверхности из-за восколипидной мембраны.)2. Облигатные анаэробные бактерии собираются в нижней части, чтобы избежать кислорода (либо не дают роста).3. Факультативные бактерии собираются в основном в верхнем (окислительное фосфорилирование является наиболее выгодным, чем гликолиз), однако они могут быть найдены на всем протяжении среды, так как от O2 не зависят.4. Микроаэрофилы собираются в верхней части пробирки, но их оптимум — малая концентрация кислорода.5. Аэротолерантные анаэробы не реагируют на концентрации кислорода и равномерно распределяются по пробирке.Анаэробы — организмы, получающие энергию при отсутствии доступа кислорода путем субстратного фосфорилирования, конечные продукты неполного окисления субстрата при этом могут быть окислены с получением большего количества энергии в виде АТФ в присутствии конечного акцептора протонов организмами, осуществляющими окислительное фосфорилирование.
Анаэробы — обширная группа организмов, как микро-, так и макроуровня:
Помимо этого анаэробное окисление глюкозы играет важную роль в работе поперечно-полосатой мускулатуры животных и человека (особенно в состоянии тканевой гипоксии).
Термин «анаэробы» ввел Луи Пастер, открывший в 1861 году бактерии маслянокислого брожения.
Анаэробное дыхание — совокупность биохимических реакций, протекающих в клетках живых организмов при использовании в качестве конечного акцептора протонов не кислорода, а других веществ (например, нитратов) и относится к процессам энергетического обмена (катаболизм, диссимиляция), которые характеризуются окислением углеводов, липидов и аминокислот до низкомолекулярных соединений.Степень аэробности среды
Интерполяция руководства к системам BD Gaspak, описывающая условия среды генерируемые пакетом[1]Для измерения потенциала среды М. Кларк предложил использовать величину ph30 — отрицательный логарифм парциального давления газообразного водорода. Диапазон [0-42,6] характеризует все степени насыщения водного раствора водородом и кислородом. Аэробы растут при более высоком потенциале [14-20], факультативные анаэробы [0-20], а облигатные — при наиболее низком [0-10].[2]
Классификация анаэробов
Согласно устоявшейся в микробиологии классификации, различают:
- Факультативные анаэробы
- Капнеистические анаэробы и микроаэрофилы
- Аэротолерантные анаэробы
- Умеренно-строгие анаэробы
- Облигатные анаэробы
Если организм способен переключаться с одного метаболического пути на другой (например, с анаэробного дыхания на аэробное и обратно), то его условно относят к факультативным анаэробам[3].
До 1991 года в микробиологии выделяли класс капнеистических анаэробов, требовавших пониженной концентрации кислорода и повышенной концентрации углекислоты (Бруцеллы бычьего типа — B. abortus)[2]
Умеренно-строгий анаэробный организм выживает в среде с молекулярным O2, однако не размножается. Микроаэрофилы способны выживать и размножаться в среде с низким парциальным давлением O2.
Если организм не способен «переключиться» с анаэробного типа дыхания на аэробный, но не гибнет в присутствии молекулярного кислорода, то он относится к группе аэротолерантных анаэробов. Например, молочнокислые и многие маслянокислые бактерии
Облигатные анаэробы в присутствии молекулярного кислорода O2 гибнут — например, представители рода бактерий и архей: Bacteroides, Fusobacterium, Butyrivibrio, Methanobacterium). Такие анаэробы постоянно живут в лишенной кислорода среде. К облигатным анаэробам относятся некоторые бактерии, дрожжи, жгутиковые и инфузории.
Токсичность кислорода и его форм для анаэробных организмов
Среда с содержанием кислорода является агрессивной по отношению к органическим формам жизни. Это связано с образованием активных форм кислорода в процессе жизнедеятельности или под действием различных форм ионизирующего излучения, значительно более токсичных, чем молекулярный кислород O2. Фактор, определяющий жизнеспособность организма в среде кислорода[4] — наличие у него функциональной антиоксидантной системы, способной к элиминации:супероксид-аниона(O2−),перекиси водорода(h3O2), синглетного кислорода(O.), а также молекулярного кислорода (O2) из внутренней среды организма. Наиболее часто подобная защита обеспечивается одним или несколькими ферментами:
- супероксиддисмутаза, элиминирующая супероксид-анион(O2−) без энергетической выгоды для организма
- каталаза, элиминирующая перекись водорода(h3O2) без энергетической выгоды для организма
- цитохром- фермент, отвечающий за перенос электронов от NAD•H к O2. Этот процесс обеспечивает существенную энергетическую выгоду организму.
Аэробные организмы содержат чаще всего три цитохрома, факультативные анаэробы — один или два, облигатные анаэробы не содержат цитохромов.
Анаэробные микроорганизмы могут активно воздействовать на среду[2] , создавая подходящий окислительно-восстановительный потенциал среды (напр. Cl.perfringens). Некоторые засеянные культуры анаэробных микроорганизмов, прежде чем начать размножаться, снижают ph30 с величины [20-25] до [1-5], ограждая себя восстановительным барьером, другие — аэротолерантные — в процессе жизнедеятельности продуцируют перекись водорода, повышая ph30[5].
Дополнительная антиоксидантная защита может обеспечиваться синтезом или накоплением низкомолекулярных антиоксидантов: витамина С, А, E, лимонной и других кислот.
Получение энергии путем субстратного фосфорилирования. Брожение. Гниение.
Схема гликолиза с образованием молочной кислоты- Также анаэробные организмы могут получать энергию путем катаболизма аминокислот и их соединений (пептидов, белков). Такие процессы именуют гниением, а микрофлору в энергетическом обмене которой преобладают процессы катаболизма аминокислот называют гнилостной.
- Анаэробные микроорганизмы расщепляют гексозы (например, глюкозу) разными путями:
- Гликолиз (Путь Эмдена-Мейергофа) после которого продукт подвергается брожению
- окислительный пентозофосфатный путь (другие названия: Фосфогликонатный путь, иначе гексозомонофосфатный(ГКМ), иначе путь Варбурга — Диккенса — Хореккера)
- Путь Энтнера — Дудорова (особенно значимый, когда субстратами служат глюконовая, маннановая, гексуроновые кислоты или их производные)
В качестве примера организма, сбраживающего сахара по пути Энтнера — Дудорова, можно привести облигатно анаэробную бактерию Zymomonas mobilis. Однако ее изучение позволяет предполагать, что Z. mobilis — вторичный анаэроб, произошедший от цитохромсодержащих аэробов. Путь Энтнера — Дудорова обнаружен и у некоторых клостридиев, что еще раз подчеркивает неоднородность эубактерий, объединенных в эту таксономическую группу.[6].
При этом характерным только для анаэробов является гликолиз, который в зависимости от конечных продуктов реакции разделяют на несколько типов брожению:
В результате расщепления глюкозы расходуется 2 молекулы, а синтезируется 4 молекулы АТФ. Таким образом общий выход АТФ составляет 2 молекулы АТФ и 2 молекулы НАД·Н2. Полученный в ходе реакции пируват утилизируется клеткой по-разному в зависимости от того, какому типу брожения она следует.
Антагонизм брожения и гниения
В процессе эволюции сформировался и закрепился биологический антагонизм бродильной и гнилостной микрофлоры:
Расщепление микроорганизмами углеводов сопровождается значительным снижением pH среды, в то время как расщепление белков и аминокислот — повышением (защелачиванием). Приспособление каждого из организмов к определенной реакции среды играет важнейшую роль в природе и жизни человека, например, благодаря бродильным процессам предотвращается загнивание силоса, заквашенных овощей, молочных продуктов.
Культивирование анаэробных организмов
Выделение чистой культуры анаэробов схематичноКультивирование анаэробных организмов в основном является задачей микробиологии.
Сложнее дело обстоит с культивированием анаэробных многоклеточных организмов, поскольку для их культивирования часто необходима специфическая микрофлора, а также определённые концентрации метаболитов. Применяется, например, при исследовании паразитов человеческого организма.
Для культивирования анаэробов применяют особые методы, сущность которых заключается в удалении воздуха или замены его специализированной газовой смесью (или инертными газами) в герметизированных термостатах — анаэростатах[7].
Другим способом выращивания анаэробов(чаще всего микроорганизмов) на питательных средах — добавление содержащих редуцирующие вещества (глюкозу, муравьинокислый натрий и др.), уменьшающие окислительно-восстановительный потенциал.
Общие питательные среды для анаэробных организмов
Для общей среды Вильсона — Блера базой является агар-агар с добавлением глюкозы, сульфита натрия и двуххлористого железа. Клостридии образуют на этой среде колонии чёрного цвета за счет восстановления сульфита до сульфид — аниона, который соединяясь с катионами железа (II) дает соль чёрного цвета. Как правило, черные на этой среде образования колонии, появляются в глубине агарового столбика.[8]
Среда Китта — Тароцци состоит из мясопептонного бульона, 0,5% глюкозы и кусочков печени или мясного фарша для поглощения кислорода из среды. Перед посевом среду прогревают на кипящей водяной бане в течение 20 — 30 минут для удаления воздуха из среды. После посева питательную среду сразу заливают слоем парафина или вазелинового масла для изоляции от доступа кислорода.
Общие методы культивирования для анаэробных организмов
GasPak — система химическим путем обеспечивает постоянство газовой смеси, приемлемой для роста большинства анаэробных микроорганизмов. В герметичном контейнере, в результате реакции воды с таблетками боргидрида натрия и бикарбоната натрия образуется водород и диоксид углерода. Водород затем реагирует с кислородом газовой смеси на палладиевом катализаторе с образованием воды, уже вторично вступающей в реакцию гидролиза боргидрида.
Данный метод был предложен Брюером и Олгаером в 1965 году. Разработчики представили одноразовый пакет, генерирующий водород, который был позднее усовершенствован ими до саше, генерирующих двуокись углерода и содержащих внутренний катализатор[9][10].
Метод Цейсслера применяется для выделения чистых культур спорообразующих анаэробов. Для этого производят посев на среду Китт-Тароцци, прогревают 20 мин при 80 °C (для уничтожения вегетативной формы), заливают среду вазелиновым маслом и инкубируют 24 ч в термостате. Затем производят посев на сахарно-кровяной агар для получения чистых культур. После 24-часового культивирования интересующие колонии изучаются — их пересеивают на среду Китт-Тароцци (с последующим контролем чистоты выделенной культуры).
Метод ФортнераМетод Фортнера — посевы производят на чашку Петри с утолщенным слоем среды, разделённым пополам узкой канавкой, вырезанной в агаре. Одну половину засевают культуру аэробных бактерий, на другую — анаэробных. Края чашки заливают парафином и инкубируют в термостате. Первоначально наблюдают рост аэробной микрофлоры, а затем (после поглощения кислорода) — рост аэробной резко прекращается и начинается рост анаэробной.
Метод Вейнберга используется для получения чистых культур облигатных анаэробов. Культуры, выращенные на среде Китта-Тароцци, переносят в сахарный бульон. Затем одноразовой пастеровской пипеткой материал переносят в узкие пробирки (трубки Виньяля) с сахарным мясо-пептонным агаром, погружая пипетку до дна пробирки. Засеянные пробирки быстро охлаждают, что позволяет фиксировать бактериальный материал в толще затвердевшего агара. Пробирки инкубируют в термостате, а затем изучают выросшие колонии. При обнаружении интересующей колонии на её месте делают распил, материал быстро отбирают и засеивают на среду Китта-Тароцци (с последующим контролем чистоты выделенной культуры).
Метод ПеретцаМетод Перетца — в расплавленный и охлаждённый сахарный агар-агар вносят культуру бактерий и заливают под стекло, помещённое на пробковых палочках(или фрагментах спичек) в чашку Петри. Метод наименее надежен из всех, но достаточно прост в применении.
Дифференциально — диагностические питательные среды
- Среды Гисса («пестрый ряд»)
- Среда Ресселя (Рассела)
- Среда Эндо
- Среда Плоскирева или бактоагар «Ж»
- Висмут-сульфитный агар
Среды Гисса: К 1 % пептонной воде добавляют 0,5 % раствор определенного углевода (глюкоза, лактоза, мальтоза, маннит, сахароза и др.) и кислотно-щелочной индикатор Андреде, разливают по пробиркам, в которые помещают поплавок для улавливания газообразных продуктов, образующихся при разложении углеводородов.
Среда Ресселя (Рассела) применяется для изучения биохимических свойств энтеробактерий(шигелл, сальмонелл). Содержит питательный агар-агар, лактозу, глюкозу и индикатор (бромтимоловый синий). Цвет среды травянисто-зелёный. Обычно готовят в пробирках по 5 мл со скошенной поверхностью. Посев осуществляют уколом в глубину столбика и штрихом по скошенной поверхности.
Среда Эндо
Среда Плоскирева (бактоагар Ж) — дифференциально-диагностическая и селективная среда, поскольку подавляет рост многих микроорганизмов, и способствует росту патогенных бактерий (возбудителей брюшного тифа, паратифов, дизентерии). Лактозоотрицательные бактерии образуют на этой среде бесцветные колонии, а лактозоположительные — красные. В составе среды — агар, лактоза, бриллиантовый зелёный, соли желчных кислот, минеральные соли, индикатор (нейтральный красный).
Висмут-сульфитный агар предназначен для выделения сальмонелл в чистом виде из инфицированного материала. Содержит триптический гидролизат, глюкозу, факторы роста сальмонелл, бриллиантовый зелёный и агар. Дифференциальные свойства среды основаны на способности сальмонелл продуцировать сероводород, на их устойчивости к присутствию сульфида, бриллиантового зелёного и лимоннокислого висмута. Маркируются колонии в чёрный цвет сернистого висмута (методика схожа со средой Вильсона — Блера).
Метаболизм анаэробных организмов
Метаболизм анаэробных организмов имеет несколько различных подгрупп:
Анаэробный энергетический обмен в тканях человека и животных[12]
Анаэробное и аэробное энергообразование в тканях человекаНекоторые ткани животных и человека отличаются повышенной устойчивостью к гипоксии (особенно мышечная ткань). В обычных условиях синтез АТФ идет аэробным путем, а при напряженной мышечной деятельности, когда доставка кислорода к мышцам затруднена, в состоянии гипоксии, а также при воспалительных реакциях в тканях доминируют анаэробные механизмы регенерации АТФ. В скелетных мышцах выявлены 3 вида анаэробных и только один аэробный путь регенерации АТФ.
3 вида анаэробного пути синтеза АТФК анаэробным относятся:
- Креатинфосфатазный (фосфогеный или алактатный) механизм — перефосфорилирование между креатинфосфатом и АДФ
- Миокиназный — синтез (иначе ресинтез) АТФ при реакции трансфосфорилирования 2 молекул АДФ(аденилатциклаза)
- Гликолитический — анаэробное расщепление глюкозы крови или запаса гликогена, заканчивающийся образованием молочной кислоты (иначе именуется «лактатным»).
Необходимо отметить, что прямым следствием гликолиза является критическое снижение рН тканей — ацидоз. Это ведет к снижению эффективного транспорта кислорода гемоглобином, и формирует положительную обратную связь.
Каждый механизм имеет свое время удержания максимальной мощности и оптимум энергообеспечения тканей. Наибольшая мощность и наименьшее время удержания:
- креатинфосфаткиназный механизм (3600 Дж/(кг·мин), при времени 6—12 сек)
- лактатный (2510 Дж/(кг·мин), при времени 30—60 сек)
- аэробный (600 Дж/(кг·мин), при времени около 600 секунд).
Примечания
- ↑ Газогенерирующие контейнерные системы GasPak: Инструкция МК. — OOO "МК, официальный дистрибьютер Becton Dickinson International", 2010. — С. 7.
- ↑ 1 2 3 К. Д. Пяткин. Микробиология с вирусологией и иммунологией. — М:"Медицина", 1971. — С. 56.
- ↑ Л. Б. Борисов. Медицинская микробиология, вирусология и иммунология. — МИА, 2005. — С. 154-156. — ISBN 5-89481-278-X
- ↑ Д. Г. Кнорре. Биологическая химия:Учеб. для хим., биол. и мед.спец.вузов. — 3. — М.:Высшая школа, 2000. — С. 134. — ISBN 5-06-003720-7
- ↑ D. A. Eschenbach, P. R. Davick, B. L. Williams. Prevalence of hydrogen peroxide-producing Lactobacillus species in normal women and women with bacterial vaginosis. — J Clin Microbiol. 1989 February; 27(2): 251–256.
- ↑ М. В. Гусев, Л. А. Минеева. Микробиология. — М:МГУ, 1992. — С. 56.
- ↑ А. А. Воробьев. Атлас по медицинской микробиологии, вирусологии и иммунологии. — МИА, 2003. — С. 44. — ISBN 5-89481-136-8
- ↑ Л. Б. Борисов. Руководство к лабораторным занятиям по медицинской микробиологии, вирусологии и иммунологии. — Медицина, 1992. — С. 31-44. — ISBN 5-2225-00897-6
- ↑ J. H. Brewer, D. L. Allgeier. Disposable hydrogen generator. — Science 147:1033-1034. — 1966.
- ↑ J. H. Brewer, D. L. Allgeier. Safe self-contained carbon dioxide-hydrogen anaerobic system. — Appl. Microbiol.16:848-850. — 1966.
- ↑ G. F. Smirnova. Metabolism peculiarities of bacteria restoring chlorates and perchlorates. — Microbiol Z. 2010 Jul-Aug;72(4):22-8.
- ↑ Филиппович Ю. Б., Коничев А. С., Севастьянова Г. А. Биохимические основы жизнедеятельности организма человека. — Владос, 2005. — С. 302. — ISBN 5-691-00505-7
См. также
Ссылки
dic.academic.ru
Анаэробы
АНАЭРОБЫ — организмы, способные жить и развиваться в бескислородной среды.[ ...]
Анаэробы организмы, живущие при отсутствии свободного кислорода.[ ...]
Анаэробы (облигатные анаэробы) — организмы, неспособные жить в кислородной среде (некоторые бактерии).[ ...]
Анаэробы - организмы, способные жить в бескислородной среде. К числу анаэробов относятся многие виды бактерий (в том числе мета-ногены).[ ...]
Анаэроб - организм, способный жить в бескислородной среде. Различают облигатных анаэробов - постоянно живущих в бескислородной среде и факультативных - способных жить как без кислорода, так и при его присутствии (организмы городской канализации, первичных отстойников и т.д.).[ ...]
Анаэробы - организмы, способные жить при отсутствии свободно! и кислорода.[ ...]
Среди проактиномицетов анаэробы встречаются чаще, чем среди актиномицетов, они обнаруживаются в организме животных и человека.[ ...]
Зеленые бактерии — строгие анаэробы и облигатные фототрофы. Исключением являются представители рода Chloroflexis. Они растут только в аэробных условиях, причем и при освещении и в темноте. Однако даже фототроф-ные бактерии, хорошо растущие в темноте, лучше развиваются при наличии света. В зависимости от организма оптимальные условия освещения для его роста могут быть различны. Одни виды хорошо растут при слабом освещении (100—300 лк), другие — при более сильном свете (700—2000 лк).[ ...]
| Рост аэробов (/), факультативных анаэробов (2) и анаэробов (3) при посеве уколом |  |
У облигатных аэробов и факультативных анаэробов в присутствии кислорода катаболизм протекает в три этапа: подготовительный, бескислородный и кислородный. В результате органические вещества распадаются до неорганических соединений. У облигатных анаэробов и факультативных анаэробов при недостатке кислорода катаболизм протекает в два первых этапа: подготовительный и бескислородный. В результате образуются промежуточные органические соединения еще богатые энергией.[ ...]
Первые микробы, по мнению Опарина, были анаэробами, так как кислорода на поверхности планеты еще не было. Затем, с появлением в составе атмосферы кислорода, часть анаэробов стала аэробами. По типу питания древнейшими микроорганизмами следует считать автотрофов. Затем, с появлением на земле растений и животных создались условия для гетеротро-фов. Сначала они вели сапрофитный образ жизни, т. е. использовали в качестве источника питания - неживые органические вещества. Затем часть сапрофитов приспособилась к жизни на поверхности и внутри других организмов, растительных и животных. Так появились паратрофы или паразиты. Что касается вирусов, то, несмотря на то, что они организованы более примитивно, чем микробы, вирусологи считают, что вирусы появились намного позже. Простота вирусов носит вторичный характер, т. е. она возникла в результате упрощения вследствие паразитического питания.[ ...]
| Содержание сульфитредуцирующих анаэробов в воде водоема в летний период. | 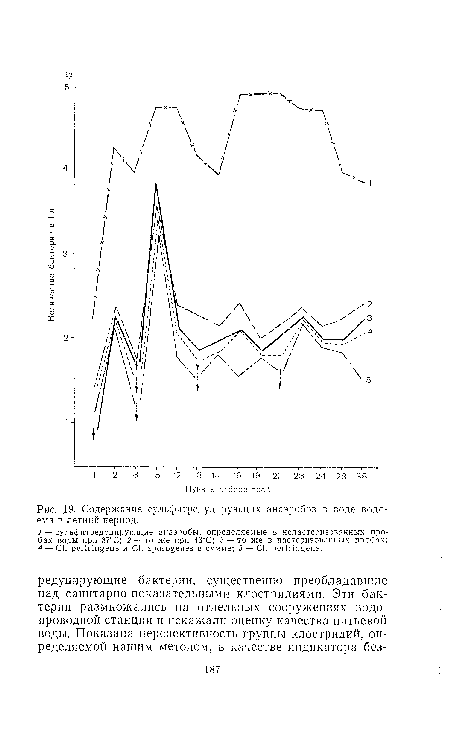 |
Большинство пурпурных серобактерий — строгие анаэробы и облигатные фототрофы, т. е. рост их возможен только при освещении. Известно лишь три вида, растущие в присутствии воздуха, причем не только на свету, но и в темноте, хотя и медленно. Это — A. roseus, E. shaposhnikovii и Т. roseopersicina. Все несерные пурпурные бактерии также растут в анаэробных условиях, но в основном являются факультативными аэробами. До недавнего времени считали, что рост пурпурных бактерий в темноте возможен лишь в аэробных или микроаэрофильных условиях, так как в отсутствие света они получают энергию в процессе дыхания. Однако недавно установлено, что R. rubrum и ряд представителей Rhodopseudo-monas растут в темноте и в строго анаэробных условиях за счет сбраживания некоторых органических субстратов. Такую же возможность, видимо, имеют пурпурные серобактерии E. shaposhnikovii и Т. roseopersicina.[ ...]
Многие из анаэробных бактерий являются «строгими анаэробами», т. е. вообще не переносят присутствия кислорода, как, например метаногенные бактерии1.[ ...]
Выросты на спорах. Замечательной особенностью спорообразования у анаэробов является формирование на спорах особых выростов с поразительно разнообразной структурой. Каждому виду анаэробных бактерий свойствен свой тип строения выростов (рис. 59). Этот признак строго специфичен, наследственно закреплен и очень устойчив. Даже у дефектных спор, потерявших способность образовывать оболочку, выросты сохраняются и не меняют своей специфической структуры.[ ...]
Значительное число бактерий — облигатных аэробов и факультативных анаэробов — способно существовать за счет использования загрязнений (примесей) воды в качестве источника питания. При этом часть использованных органических веществ расходуется на энергетические нужды, а другая часть — на синтез тела клетки. Часть вещества, расходуемая на энергетические потребности, окисляется клеткой до конца, т. е. до СОг, НгО, >Ш3. Продукты окисления -— метаболита — выводятся из клетки во внешнюю среду. Реакции синтеза клеточного вещества идут также с участием кислорода. Количество кислорода, требуемого микроорганизмам на весь цикл реакции синтеза и получения энергии, и есть БПК.[ ...]
Токсическое действие кислорода воздуха на рост и развитие облигатных анаэробов и тяготение к низкому окислительно-восстановительному потенциалу, по современным представлениям, можно объяснить тем, что молекулярный кислород и высокий ОВП могут служить причиной необратимого окисления жизненно важных ферментов, обусловливающих основные процессы их метаболизма.[ ...]
Таким образом, при дыхании конечным акцептором водорода является кислород. У анаэробов в качестве акцепторов водорода выступают либо органические субстраты (брожение), либо неорганические вещества, такие, как нитраты или сульфаты («анаэробное дыхание»). Из схемы видно, что наиболее просто и примитивно транспорт электронов осуществляется у большинства анаэробов из-за отсутствия у них ферментов цепи переноса электронов, способных передавать электроны по цепочке вплоть до молекулярного кислорода.[ ...]
К строго автэтрофным организмам относятся аэробы Thiobacillus thioparus и Thiobacillus thiooxidans и анаэроб Thiobacillus thiocyanooxidans, который был выделен в 1954 г. Хапполдом и Джонстоном из сточных од газового завода, содержащих тиоционат. Эта культура бактерий, разрушающая тиоционат с образованием аммиака и сульфатов, способна окислять также тиосульфат и серу.[ ...]
Метанообразующие бактерии Methano bacterium omelianskii, Bad. formicicum, Methanosarcina barkeri являются облигатными анаэробами и относительно плохо поддаются выделению. Культура Bad. formicicum разлагает муравьиную кислоту с образованием различных продуктов распада, причем направление процесса зависит от окислительно-восстановительного потенциала среды. В условиях относительного анаэробиоза, как установил JI. В. Омелянский [10 ]j муравьиная кислота разлагается с образованием водорода и углекислого газа; при этом потенциал питательной среды снижается до гН2 12—12,9 и создаются анаэробные условия. При разложении в анаэробных условиях и снижении гН2 до 6—7 муравьиная кислота разлагается с образованием метана; в интервале значений гН2 16—22 разложение муравьиной кислоты идет лишь с образованием углекислого газа.[ ...]
Терин [493] выделил из содержимого метантенков и охарактеризовал 92 культуры. Все бактерии были анаэробами или микро-аэрофилами. Около 50% изолятов были спорообразующими. Палочковидные микроорганизмы напоминали по морфологии Согу-nebacterium, Lactobacillus, Ramibacterium, Actinomyces и Bifidobacterium. Автор считает, что классическое определение до вида бактерий, осуществляющих анаэробное разложение органических отходов, меньше дает для характеристики бактериологии процесса, чем выделение крупных микробных групп на основании морфологического и биохимического сходства.[ ...]
Род Staphylococcus. Представители этого рода отличаются от микрококков тем, что являются факультативными анаэробами и вызывают ферментацию глюкозы в анаэробных условиях с образованием кислоты, кроме того, многие виды стафилококков патогенны. Большинство видов требуют органических источников азота и витаминов для роста. Многие штаммы образуют оранжевый или желтый пигмент. Типовой вид рода — Staphylococcus aureus.[ ...]
Энергетический обмен. По отношению к свободному кислороду организмы делятся на три группы: аэробы, анаэробы и факультативные формы.[ ...]
Род Acidianus - сферические клетки, иногда дольчатой формы и выглядят как тетраэдры, пирамиды и т.п. Факультативные анаэробы: в аэробных условиях они окисляют серу и Fe2+, в анаэробных - восстанавливают серу молекулярным водородом.[ ...]
Из полученных промежуточных продуктов распада кислой фазы в щелочной фазе образуются метан и углекислота. Эту фазу брожения осуществляют анаэробы — метанообразующие бактерии.[ ...]
Из микроорганизмов на кормах часто развиваются Вас. Возбудитель ботулизма широко распространен в природе. Спорообразующий почвенный анаэроб развивается в загрязненных землей, птичьим пометом или испорченных кормах и выделяет весьма сильные и стойкие токсины. Особенно благоприятные субстраты для этого микроба - полова или мякина и влажное зерно, а также подвергающиеся самосогреванию, неправильно заскирдованные солома и сено, а также силос, загрязненный частицами земли и трупами грызунов.[ ...]
Некоторые микроорганизмы могут жить как в присутствии свободного кислорода воздуха, так и в отсутствие его. Они относятся к факультативным, или условным, анаэробам. Примером факультативных анаэробов являются молочнокислые бактерии, дрожжи и др.[ ...]
Большинство лучистых грибков развивается при свободном притоке кислорода. Однако отдельные культуры могут расти и при некотором ограничении доступа кислорода. Строгие анаэробы среди лучистых грибков встречаются редко.[ ...]
Главным условием для проведения точных экспериментов по физиологии, биохимии, цитологии и генетике микроорганизмов является выделение чистых культур. Получить чистую культуру анаэробов очень сложно. В последнее время были созданы специальные приборы — анаэростаты, в которых микроорганизмы можно культивировать на плотных питательных средах в атмосфере, почти полностью лишенной кислорода. Но и при использовании современных приборов и средств выделение и инкубирование чистых культур с высокой степенью анаэробности является крайне трудным делом. Поэтому, вероятно, подавляющее большинство, например, сульфатредуцирующих, цел-люлозолитических и других спороносных анаэробов остаются до сих пор неизвестными. По существу, из этих физиологических групп, широко распространенных в природе, в лабораториях получены лишь единичные культуры.[ ...]
Первая культура анаэробных спорообразующих бактерий, способная осуществлять брожение целлюлозы, была выделена В. Л. Оме-л янским. В литературе описано 12 видов целлюлозолитических анаэробов. Спорообразукь щие целлюлозолитические анаэробы до настоящего времени очень плохо изучены в связи с трудностями их выделения и культивирования. Большинство выделенных штаммов были загрязнены посторонними бактериями, что значительно обесценивает данные по физиологии и биохимии этих организмов.[ ...]
По характеру диссимиляции различают аэробные и анаэробные организмы. Аэробные (от греч. аег — воздух) организмы для дыхания (окисления) используют свободный кислород. Аэробами является большинство ныне живущих организмов. Напротив, анаэробы окисляют субстраты, например, сахара в отсутствие кислорода, следовательно, для них дыханием является брожение. Анаэробами являются многие микроорганизмы, гельминты. Например, динитрифицирующие анаэробные бактерии окисляют органические соединения, используя нитриты, являющиеся неорганическим окислителем.[ ...]
Но известны животные, которые могут одинаково нормально жить и при хорошей доступности кислорода, и при крайне низком содержании его, и почти при полном отсутствии, и даже такие, которым кислород не только не нужен, но даже вреден. Первых называют анаэробами факультативными, вторых — облигатными. К первым относятся водные черепахи и многие рыбы, ведущие придонный образ жизни. Дело в том, что в придонной воде содержание кислорода может доходить до 15 % той величины, которая наблюдается при насыщении воды воздухом.[ ...]
Приведенные результаты позволяют заключить, что ведущую роль в процессах анаэробного разложения органического материала играют облигатные анаэробные бактерии. Однако систематическое выявление в содержимом метантенков аэробов и факультативных анаэробов свидетельствуют о том, что эти микроорганизмы также участвуют в деструкции органических веществ, и при определенных условиях численность их может существенно возрастать. Так, при добавлении к ферментируемой жидкости глюкозы количество аэробных и факультативно анаэробных бактерий повышается от 1 X 10б до 3,2 X 109 клеток/мл (цит. по [404]).[ ...]
При нормальной работе искусственных аэробных очистных сооружений основная масса бактерий состоит из аэробов — микробов, нуждающихся для своей жизнедеятельности в наличии молекулярного атмосферного кислорода, и лишь в незначительном количестве встречаются факультативные анаэробы — организмы, способные развиваться как в присутствии кислорода, так и в его отсутствии. Некоторые аэробы могут использовать не только кислород атмосферы, но и кислород минеральных соединений (например, нитратов, сульфатов).[ ...]
В первой фазе сбраживания, которая называется кислой или водородной, из углеводов, жиров и белков образуются основные продукты распада — жирные кислоты, водород, углекислый газ, спирты аминокислоты, аммиак, сероводород и др. Бактерии, осуществляющие первую фазу сбраживания осадка, являются факультативными анаэробами типа дрожжей, маслянокислых, пропионовых, бутиловых и др.[ ...]
Присутствие кислот в среде обусловливает ее кислую реакцию. Кроме НЖК продуктами распада первой фазы являются низшие жирные спирты, аминокислоты, некоторые альдегиды и кетоны, глицерин, а также углекислота, водород, сероводород, аммиак и некоторые другие соединения. Осуществляют эту фазу процесса бактерии, относящиеся к факультативным анаэробам (молочнокислые бактерии, уксусно-кислые, пропионово-кислые и т. п.) и к облигатным анаэробам (масляно-кислые бактерии, целлюлозные, ацетонобутиловые и др.).[ ...]
Таким образом, превращение органического вещества в ме-тантенках происходит в две стадии: сбраживание субстрата до жирных кислот (неметаногенная) и образование из жирных кислот СН4 и С02 (метаногенная). Во время первой стадии главную роль играют анаэробные бактерии родов Clostridium, Bacteroides и др. Вторую стадию осуществляет уникальная группа облигатных анаэробов — метановые бактерии родов Methano-bacterium, Methanobacillus, Methanococcus, Methanosarcina.[ ...]
На этой точке зрения особенно настаивает Маргулис (Margulis, 1975). Позднее поглощение клеток сине-зеленых водорослей могло бы привести к появлению автотрофного «организма», предка всего царства растений. Эта последовательность событий является чисто гипотетической и вызывает много скептических замечаний, но, по крайней мере отчасти, она может соответствовать действительности. До сих пор существуют близкие к предполагаемым симбионтам формы, включая бактерию Paracoccus denitrificans, рядом признаков напоминающую гипотетического свободножи-вущего предшественника митохондрии. Если теория Маргулис верна, то основная часть этой главы была посвящена второй стадии развития мутуализма, когда все более интегрируются друг с другом пары видов, каждый из которых по происхождению является симбиотической ассоциацией.[ ...]
Анаэробиоз некоторые авторы объясняют тем, что эти микробы не имеют фермента каталазы, разлагающего перекись водорода. Считают, что на первой стадии процесса окисления органического вещества кислородом воздуха образуется перекись водорода, которая является ядом для всех живых существ, но все микробы, кроме анаэробных, способны выделять в среду фермент каталазу, разлагающий перекись водорода на воду и кислород. Анаэробы в присутствии кислорода отравляются перекисью водорода.[ ...]
Один из важнейших физико-химических показателей осадков, определяющих рост и развитие некоторых групп микроорганизмов, -окислительно-восстановительный ЕЬ-потенциал. Он отражает также меру аэробности системы, что особенно важно при характеристике осадков сточных вод. Бактерии классифицируют по зонам потенциала. Бактерии более активны при снижении ЕЬ-потенциала с 400 до 200 мВ. Анаэробные бактерии функционируют лучше при изменении ЕЬ-потен-циала с 50 до 400 мВ. Факультативные анаэробы функционируют в обеих зонах потенциала, но активность их повышается в зоне отрицательного потенциала.[ ...]
Формирование микрофлоры метантенка происходит за счет микроорганизмов, попавших вместе со сточными водами или осадком. По видовому составу биоценоз метантенков значительно беднее аэробных биоценозов, из них выделено лишь около 50 видов бактерий, способных осуществлять первую стадию расщепления загрязнений - стадию кислотообразования. Наряду с облигатными анаэробами в метантенке могут встречаться и факультативные анаэробы. Общее количество бактерий в осадке колеблется от 1 до 15 млг/мл. Конечным продуктом процесса брожения этой группы микроорганизмов являются низшие жирные кислоты, диоксид углерода, ионы аммония, сероводород .[ ...]
Ингибирующее действие Lactobacillus spp. на другие микроорганизмы обусловлено их способностью вырабатывать кислоту. Именно эта молочная кислота поддерживает кислую реакцию среды в кишечнике большинства животных. Многие энтеро-патогенные микробы предпочитают нейтральную или слегка щелочную среду, так что сама по себе кислотность может угнетать их. Кроме того, L. acidophilus является факультативным анаэробом и бурно размножается в кишечнике. При таких условиях лактобациллы имеют заметное преимущество перед другими микроорганизмами в борьбе за питательные вещества.[ ...]
Жизнь в толще литосферы существует в основном в подземных водах.[ ...]
Для ускорения анализа перспективна возможность оденивать результат по появлению и накоплению промежуточных, а в некоторых „случаях и конечных продуктов разложения составных частей питательных и диагностических сред. Наличие и размножение микроорганизмов можно оценивать после макро- или микрохимического определения продуктов их метаболизма при соответствующем составе питательных сред. Примером в этом отношении может служить среда Вильсона —■ Блера с сульфитом натрия и хлоридом железа для обнаружения суль-фитредуцирующих анаэробов. При наличии в посеве анаэробов сферическая черная зона из сернистого железа образуется уже к 8-му часу, т. е. значительно раньше, чем накопится масса клеток для образования видимых колоний.[ ...]
В нашей стране в конце 40-х годов также было начато изучение антибиотических свойств лишайников. В результате этих исследований в Ботаническом институте АН СССР в Ленинграде был получен новый медицинский препарат — натриевая соль усниновой кислоты, или «Бинан». Основой для получения препарата явилась усниновая кислота. Исследование антимикробных свойств препарата показало, что он активен против грамположи-тельной бактериальной флоры — золотистого стафилококка, различных стрептококков, пневмококков, анаэробов и туберкулезной палочки. Препарат представляет собой эффективное наружное антимикробное средство для лечения нагноительных процессов в раневых поверхностях. В настоящее время это лекарство широко продается в аптеках в нескольких формах: в водно-спиртовых новокаиновых растворах, в касторовом масле с анестезином, в пихтовом бальзаме и в виде порошка. Препарат «Бинан» нашел применение в хирургической практике при лечении свежих посттравматических и послеоперационных раневых поверхностей, при лечении варикозных и трофических язв, острого гнойного воспалительного процесса мягких тканей, травматических остеомиелитов, при пластических операциях, при лечении ожогов II и III степени. Его применяют также в гинекологии.[ ...]
При биологическом окислении идут окислительно-восстановительные реакции, сопровождающиеся отнятием атомов водорода от одних соединений (доноров) и передачей его другим (акцепторам), или реакции, связанные с переносом электронов от донора к акцептору. Эти процессы осуществляются при участии ферментов, относящихся к классу оксиредуктаз. Процессы дыхания, в которых акцептором водорода или электронов является молекулярный кислород, называются аэробными. Если же акцепторами будут другие неорганические или органические соединения, то такой тип дыхания называется анаэробным. По типу дыхания выделяют две группы микроорганизмов: аэробы (оксибиотические формы), которым для дыхания необходим кислород, и анаэробы (аноксибиотические формы), развивающиеся в отсутствие кислорода. Между ними нет резкого различия. Наряду со строгими (облигатными) аэробами и анаэробами есть микроорганизмы, которые могут жить в присутствии кислорода и без него. Это микроаэрофилы, оптимум содержания кислорода в воздухе для которых составляет 0,5—1%, и факультативные анаэробы. Так, кишечная палочка является факультативным анаэробом.[ ...]
Температура сбраживания является одним из важнейших факторов, влияющих на скорость распада осадка в метан-тенках. Большинство метан-тенков эксплуатируется в условиях мезофильного брожения при температуре 30—35°. При этом распад органического вещества осуществляется теми же группами микроорганизмов, которые вызывают его в двухъярусном отстойнике, работающем обычно при температуре 10—20°. Однако повышение температуры не только ускоряет процесс распада, но делает его более глубоким. При термофильном брожении (50—55°) в процессе распада принимает участие специфическая группа анаэробных термофильных микроорганизмов, имеющих весьма энергичный обмен вследствие высокой активности присущих им ферментов.[ ...]
ru-ecology.info
Аэробы строгие - Справочник химика 21
Дыхание. Большинство гетеротрофных организмов получает энергию в результате биологического окисления органических веществ — дыхания. Водород от окисляемого вещества (см. 24) передается в дыхательную цепь. Если роль конечного акцептора водорода выполняет только кислород, процесс носит название аэробного дыхания, а микроорганизмы являются строгими (облигатными) аэробами, которые обладают полной цепью ферментов переноса (см. рис. 14) и способны жить только при достаточном количестве кислорода. К аэробным микроорганизмам относятся многие виды бактерий, гри-бь1, водоросли, большинство простейших. Аэробные сапрофиты играют основную роль в процессах биохимической очистки сточных вод и самоочищении водоема. [c.63] Ввиду описанных преимуществ дыхания не удивительно, что на планете, атмосфера и гидросфера которой богаты Ог, эволюция большей части животного царства привела к абсолютной зависимости от аэробного метаболизма. Однако не у всех организмов и не у всех тканей одного и того же организма зависимость от кислорода выражена в одинаковой степени. Например, скелетные мышцы позвоночных способны работать главным образом за счет гликолиза это происходит в короткие периоды интенсивной деятельности, когда поступление кислорода оказывается недостаточным, чтобы весь образующийся пируват поступал в цикл Кребса. Мозговое вещество почки тоже может в значительной степени использовать анаэробный обмен. Некоторые другие ткани, особенно сердечная мышца и центральная нервная система, полностью зависят от дыхания. Большинство тканей занимает промежуточное положение и может хотя бы короткое время переносить анаэробиоз. Подобно тому как разные ткани одного организма могут сильно различаться по зависимости от О2. разные виды организмов также могут сильно различаться по своей зависимости от дыхания. В этом отношении одну крайнюю группу составляют строгие аэробы. Они не могут жить без кислорода. Другая крайняя группа организмов — облигатные анаэробы для выживания их необходимо полное отсутствие О2. Промежуточный класс, к которому относятся многие виды беспозвоночных, образуют факультативные анаэробы. Эти организмы используют кислород, когда он имеется, но могут неограниченно долго выживать и при полном его отсутствии. [c.44]При биологическом окислении идут окислительно-восстановительные реакции, сопровождающиеся отнятием атомов водорода от одних соединений (доноров) и передачей его другим (акцепторам), или реакции, связанные с переносом электронов от донора к акцептору. Эти процессы осуществляются при участии ферментов, относящихся к классу оксиредуктаз. Процессы дыхания, в которых акцептором водорода или электронов является молекулярный кислород, называются аэробными. Если же акцепторами будут другие неорганические или органические соединения, то такой тип дыхания называется анаэробным. По типу дыхания выделяют две группы микроорганизмов аэробы (оксибиотические формы), которым для дыхания необходим кислород, и анаэробы (аноксибиотические формы), развивающиеся в отсутствие кислорода. Между ними нет резкого различия. Наряду со строгими (облигатными) аэробами и анаэробами есть микроорганизмы, которые могут жить в присутствии кислорода и без него. Это микроаэрофилы, оптимум содержания кислорода в воздухе для которых составляет 0,5—1%, и факультативные анаэробы. Так, кишечная палочка является факультативным анаэробом. [c.216]
При глубинном культивировании микроорганизмы развиваются во всем объеме жидкой питательной среды. Так как подавляющее большинство продуцентов ферментов — строгие аэробы, среду интенсивно аэрируют. В микроорганизмах протекают два неразрывно связанных процесса — синтез биомассы и синтез ферментов. [c.159]
Отношение к Ог Строгий аэроб Строгий аэроб [c.21]
Среди живых организмов, в зависимости от характера их обмена, принято различать аэробные и анаэробные. Строгими анаэробами в истинном смысле этого слова являются только немногие группы бактерий, которые не только не способны использовать кислород, ио даже им отравляются . Огромное большинство микроорганизмов относится к факультативным (условным) анаэробам-, они используют кислород, когда последний доступен, но могут существовать и в анаэробных условиях. Животные большей частью должны быть отнесены к строгим аэробам, так как только очень немногие из них [c.374]
Строгий аэроб Нуждается в дрожжевом автолизате добавка тиосульфата стимулирует рост 6.9 2,8 [c.41]
Плесневые грибы переносят сильное подкисление, но являются строгими аэробами, поэтому в хорошо спрессованном, укрытом, заквашенном корме размножаться не мОгут. [c.194]
Группа 14. Грамположительные, не образующие спор палочки правильной формы. Группа — конгломерат, состоящий из 7 родов, объединенных несколькими общими морфологическими и физиологическими признаками клетки палочковидной формы (от кокковидных до удлиненных, одиночных или образующих цепочки), мезофилы строгие или факультативные аэробы, есть микроаэрофилы и аэротолерантные анаэробы хемоорганогетеротрофы, растущие только на сложных средах. [c.173]
Поли-р-гидроксимасляную кислоту образуют многие аэробные бактерии (рис. 1.12), а также цианобактерии и анаэробные фототрофные бактерии. У факультативных анаэробов и строгих аэробов она накапливается, когда клетки испытывают недостаток О2 и переходят на брожение поэтому ее можно рассматривать как полимерный внутриклеточный продукт брожения. В аэробных условиях этот продукт может быть снова вовлечен в метаболизм как источник энергии и углерода и использован в процессе дыхания. [c.34]
Являются строгими аэробами. [c.89]
Другим важным процессом наряду с питанием является дыхание, благодаря которому клетка получает необходимую ей энергию. Большинство микроорганизмов при дыхании используют кислород воздуха (аэробы), но есть и такие, которые растут без доступа воздуха анаэробы). Помимо строгих аэробных и анаэробных микроорганизмов, существуют необязательные, так на- [c.12]
Это диплококки бобовидной формы, в мазках имеют вид кофейных зерен. Спор не образуют, жгутиков не имеют, в организме образуют капсулу. Грамотрицательные. Строгие аэробы. [c.94]
Все эти бактерии—автотрофы (см. п. 1.3.1) и строгие аэробы. Они используют энергию окисления аммонийного азота и нитритов для восстановления неорганического углерода, источником которого являются диоксид углерода и карбонаты. [c.406]
Дыхание бактерий происходит при помощи специальных дыхательных ферментов. По отношению к молекулярному кислороду (по типу дыхания) микробы разделяются на аэробов и анаэробов. Между строгими анаэробными и аэробными типами дыхания существуют и переходные формы. [c.45]
Конечно, внутри дыхательной цепи место каждого окислительно-восстановительного соединения, если уж оно присутствует, более или менее детерминировано его стандартным потенциалом. Так, пиридиннуклеотид должен стоять в начале, флавопротеид — в середине, а цитохромы — в конце цепи. Более того, присутствие некоторых окислительно-восстановительных соединений кажется вероятным. Пиридиннуклеотид в любом случае содержится у всех бактерий, в том числе и у строгих анаэробов, а флавопротеиды обнаруживаются как у многих анаэробов, так и у всех аэробов. [c.146]
Позже многие из фотосинтезирующих бактерий, ставшие аэробными, утратили способность к фотосинтезу, превратившись в чисто дышащие организмы, т. е. в строгих аэробов среди бактерий. Утрата способности к фотосинтезу хорошо известна также среди сине-зеленых водорослей, от которых произошли бесцветные скользящие бактерии , энергетика которых основана на процессах дыхания 14,Ж). По анало- [c.149]
Миксобактерии близки к истинным бактериям, они имеют такие же малые размеры и палочковидную форму размножаются путем полеречного изоморфного деления являются хемосинтезирующими, гетеротрофами способны расти на искусственных средах. Большинство из них строгие аэробы, не образующие опор мезофнлы, растут гари нейтральной реакции среды pH 7,2. [c.281]
Хотя облигатно анаэробные бактерии в целом очень чувствительны к О2, они могут в природе находиться в аэробных зонах. Широкое распространение представителей рода lostridium в местах с высоким парциальным давлением Oj объясняется наличием у них эндоспор, не чувствительных к молекулярному кислороду. Однако и многие не образующие спор строго анаэробные прокариоты обнаружены в природе в местах, где наблюдается активное развитие облигатных аэробов. Вероятно, совместное развитие с облигатными аэробами, активно потребляющими молекулярный кислород, приводящее к образованию зон с низкой концентрацией [c.129]
К V подгруппе отнесены архебактерии, характеризующиеся совокупностью следующих признаков облигатные термофилы ацидофилы или нейтрофилы аэробы, факультативные или строгие анаэробы автотрофы или гетеротрофы. Метаболизм больщинства из них связан с молекулярной серой (S ). Представители порядка Thermoproteales в процессе хемолитоавтотрофного роста получают энергию в реакции [c.180]
По типу дыхания все бактерии можно разделить на три группы 1) строгие анаэробы — маслянокислые бактерии, возбудители ботулизма 2) строгие аэробы — уксуснокислые бактерии, палочки инфлуэнцы, псевдомонасы 3) факультативные анаэробы — кишечная палочка, молочнокислые бактерии, по-давляюшее большинство патогенных бактерий и дрожжи. У представителей всех этих трех групп возможно дыхание путем непрямого окисления — дегидрогенирования. С отщеплением водорода освобождается электрон, а энергия поступает на нужды микробной клетки. Например, некоторые водные микроорганизмы осуществляют реакцию [c.94]
По отношению к кислороду в среде, где происходит вегетация, микроорганизмы делятся на облигатные (строго обязательные) аэробы, облигатные (строго обязательные) анаэробы, факультативные анаэробы (растущие в аэробных и в анаэробных условиях), микроаэрофилъные анаэробы (переноснш,ие малое количество кислорода). [c.22]
Особенно эффективно связывают азот виды Azotoba ter (около 20 мг азота на 1 г использованного сахара). Различают несколько видов Azotoba ter, распространенных в разных местообитаниях (табл. 13.1). Все они грам-отрицательные, относительно крупные, в определенных условиях передвигаются с помощью жгутиков все-строгие аэробы, способные окислять многие органические соединения. У А. hroo o um клетки соединены попарно. Обильное образование слизи и наличие темных пигментов (меланинов) придают колониям характерный вид. При недостатке питательных веществ образуются цисты с толстыми клеточными стенками ( артроспоры , микроцисты ). [c.399]
По методу Грама мпкроб окрашивается отрицательно. Он является строгим аэробом и весьма требователен к питательным средам. Па обычных питательных средах, как бульон, картофель, мясонептонная желатина и т. д., он не растет. Эта бактерия хорошо развивается на среде из яичного желтка, сыворотке животных или их крови. [c.486]
Уникальной характеристикой активных клубеньков является присутствие красного пигмента, леггемоглобина. Подобно гемоглобину млекопитающих леггемоглобин обладает важным свойство.м — способностью к связыванию и иммобилизации кислорода. Существенно также, что этот процесс обратим. Значение этого белка, производимого бобовыми и располагающегося в цитоплазме хозяина [558], было понято только после осознания роли кислорода. Ризобактерии — строгие аэробы, они могут расти и размножаться только в присутствии кислорода. Однако энзим, который они вырабатывают, нитрогеназа, инактивируется при высоком содержании кислорода и может фиксировать азот только в бескислородной атмосфере. Похоже, что леггемоглобин осуществляет необходимую регуляцию распределения кислорода внутри клубеньков. Он связывается с молекулярным кислородом, так что нитрогеназа не ингибируется, но связанный кислород может быть доступным в дыхательных центрах в цитоплазме хозяина вблизи бактероидов [559]. Структурная организация клубеньков также способствует быстрому удалению продуктов фиксации азота, накопление которых ингибирует связывание. [c.278]
Все типы отношения к кислороду найдены среди бактерий и простейших. Грибы, в основном, аэробы, но некоторые виды, как правило, дрожжевые анаморфы — факультативные анаэробы. Есть и небольшое количество строго анаэробных видов. Водоросли — почти всегда облигатные аэробы. [c.97]
Десятая группа — скользящие бактерии, образующие плодовые тела — являются хемоорганотрофами и строгими аэробами без жгутиков, которые могут скользить по твердой поверхности. При исчерпании питательных веществ клетки агрегируют и формируют плодовые тела, состоящие из модифицированной слизи и клеток, часто ярко окрашенные и имеющие макроскопические размеры. Плодовые тела различаются по сложности строения и размерам. Внутри плодовых тел формируются миксоспоры или микроцисты (пор. Myxo o ales). [c.325]
Десятая группа — образующие эндоспоры грамположительные палочки и кокки, формируют устойчивые к нагреванию эндоспоры. Подвижны, в основном палочки или нити. Один род Sporosar ina) содержит подвижные кокки в тетрадах или кубических пакетах. Строгие аэробы, микроаэрофилы, факультативные или строгие анаэробы (представители родов Ba illus, lostridium). [c.331]
Двенадцатая группа — неправильные неспорулирующие грамположительные палочки. Окраска грамвариабельна. Растут в присутствии воздуха и не образуют эндоспор. От строгих аэробов до строгих анаэробов. Могут быть патогенны для животных и растений (во всех трех классах филума Firmi utes). [c.331]
Неправильные бесспоровые грамположительные палочки. Могут формировать разветвленные нитчатые элементы, смеси палочек и нитей, коккоидные формы. Некоторые имеют жизненный цикл палочки —кокки . От строгих анаэробов до строгих аэробов. Есть представители, патогенные для животных или растений и обычные обитатели ЖКТ и рубца (роды Arthroba ter, Bifidoba terium, Propioniba terium). [c.332]
Стрептомицеты и родственные роды. Большинство — строгие аэробы. Хемогетеротрофы. Мицелиальный рост часто сопровождается сильным развитием воздушного мицелия с длинными цепочками спор. Колонии обычно ярко пигментированы. Растут медленно, используют щирокий спектр соединений. Обладают мощными гидролазами. Клеточная стенка содержит L-ДАП и глицин. Типичные обитатели почв, есть патогенные виды (актиномико-зы). Ряд представителей является продуцентами важнейших антибиотиков (род Streptomy es). [c.333]
По другой методике к капле добавляют примерно 10 мкл стерильной среды и инкубируют на предметном стекле в закрытой камере до появления клона бактерий, а затем извлекают несколько клеток капиллярной пипеткой. Поскольку рост клона происходит в полуана-эробных условиях, эта методика неприменима для строгих аэробов. [c.324]
chem21.info





















 |  |  |  |  |  |  |  |  |  |  |  |  |  |  |
 Пример видео 3 Пример видео 3 |  Пример видео 2 Пример видео 2 |  Пример видео 6 Пример видео 6 |  Пример видео 1 Пример видео 1 |  Пример видео 5 Пример видео 5 |  Пример видео 4 Пример видео 4 |
Администрация муниципального образования «Городское поселение – г.Осташков»