



КЛАСС СУМЧАТЫЕ ГРИБЫ ИЛИ АСКОМИЦЕТЫ (ASCOMYCETES). Аскомицеты дрожжи
1. Характерные признаки Ascomycota
Мицелий многоклеточный, гаплоидный, одноядерный или многоядерный, а у некоторых дикариотический, септированный. В центре септы – пора, через которую осуществляется движение цитоплазмы и органелл, в частности, ядра. У дрожжей (Saccharomycetales) мицелий вторично упрощен, способен к почкованию. В оболочках клеток – хитин и β-глюкан. У видов класса Hemiascomycetes — маннан и β -глюкан. Клеточные стенки двуслойные с тонким наружным слоем и относительно толстым внутренним слоем. Митоз внутриядерный.
Жизненный цикл с зиготической редукцией, ядра после мейоза делятся один раз митотически, формируя эндогенные аскоспоры в аске (сумке). В сумке происходит 4 процесса: кариогамия, мейоз, митоз, дифференциация аскоспор. При половом процессе ядра не сливаются, а попарно координируются в дикарион, синхронно делятся и образуют дикариотический мицелий – аскогенные гифы. В цикле развития преобладает гаплофаза.
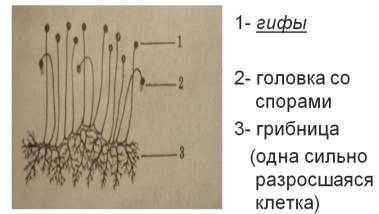
Бесполое размножение экзогенными конидиями, формирующимися на конидиеносцах разнообразного строения. Конидиеносцы как одиночные так и многочисленные, собранные в пучки или развивающиеся в особых вместилищах – ложах и пикнидах. Освобождение конидий обычно пассивное. У паразитных аскомикота конидиальное спороношение чаще развивается на живом растении-хозяине, а сумчатое – уже на его отмерших частях (листьях, стеблях).
Сумчатые грибы играют значительную роль в природных сообществах суши; лишайники (в состав которых входят аскомикота) являются основными продуцентами в тундровой зоне; многие (например, Erysiphales) – опасные паразиты сельскохозяйственных культур. Многие сумчатые продуценты антибиотиков, витаминов и ферментов.
2. Половой процесс Ascomycota
Характерный половой процесс - гаметангиогамия. Встречаются гомоталличные и гетероталличные виды.
Типичный половой процесс изучен у Руrопета omphaloides. Женский половой орган – архикарп состоит из двух многоядерных клеток: большей – аскогона и отходящей от него вверх меньшей – трихогины, быстро теряющей свое содержимое. Антеридий, одноклеточный и многоядерный, при оплодотворении переливает своё содержимое через трихогину в аскогон, т.о. происходит плазмогамия. Мужские и женские ядра не сливаются, а ассоциируются в пары – дикарионы. Аскогон образует нитевидные выросты, называемые аскогенными гифами, каждая клетка которых содержит дикарион.
Конечная клетка аскогенной гифы загибается в виде крючка, а ядра её дикариона синхронно делятся в месте перегиба (рис. 10). Одна пара ядер остаётся в месте перегиба, а вторая пара разделяется: одно ядро переходит в кончик крючка, а второе — в его основание. Затем возникают две перегородки, отделяющие две одноядерные и одну двуядерную клетку. Две одноядерные клетки сливаются (без слияния ядер), восстанавливая двуядерность базальной клетки, которая в дальнейшем способна повторить указанный цикл. Двуядерная клетка развивается в сумку. Она увеличивается в размерах, вытягивается, ядра дикариона сливаются, затем диплоидное ядро мейотически делится; после чего следует митотическое деление 4 гаплоидных ядер, вокруг которых формируются 8 аскоспор. После многократного повторения этих процессов образуется пучок сумок.
|
| Рис. 10. Половой процесс и развитие сумки у Руrопета 1 – антеридий 2 – архикарп 3 – аскогон (стадия дикариона) 4 – трихогина 5-8 – развитие сумки на аскогенных гифах 5 – кариогамия 6 – мейоз 7 – митоз 8 – формирование аскоспор |
Пучок сумок обрастает плотным сплетением гаплоидных гиф, образующих плодовое тело, в котором зрелые сумки защищены от возможных повреждений. К моменту созревания аскоспор тургорное давление в сумке резко возрастает и аскоспоры с силой выбрасываются.
Таким образом, в цикле развития большинства аскомикота чередуются гаплоидная стадия (аскоспора, мицелий, конидии, половые органы), дикариотичная стадия (оплодотворённый аскогон, аскогенные гифы) и короткая диплоидная стадия (молодая сумка с диплоидным ядром). Плазмогамия и кариогамия разделены кратковременной стадией дикариона.
У некоторых сумчатых имеются отклонения от описанного процесса.
Отсутствие стадии дикариона характерно для морфологически наиболее примитивных аскомицетов – пекарских дрожжей (Saccharomyces cerevisiae). Дрожжи сахаромицеты имеют в клетках диплоидные ядра и на богатой сахарами среде размножаются вегетативно — почкованием. При обеднении среды ядро редукционно делится, и клетка превращается в сумку, содержащую 4 гаплоидные споры. Оболочка клетки растворяется, и споры освобождаются. Если они оказываются далеко друг от друга, то начинают размножаться почкованием, если же поблизости, то под влиянием половых феромонов попарно сливаются, причем ядра сливающихся клеток сближаются и также сливаются, после чего вновь образуется диплоидная клетка.
Мицелий небольшого порядка сумчатых грибов (Taphrinales) является дикариотичным. Перед образованием сумок ядра в клетках сливаются и сразу редукционно делятся. После двух делений мейоза происходит еще одно митотическое и формируется сумка с 8 гаплоидными аскоспорами. Выходя наружу, аскоспоры способны размножаться почкованием или образовывать двухъядерные клетки (в результате слияния). Такие клетки могут заражать растения, развивая в их тканях двухъядерный мицелий (мицелиально-дрожжевой диморфизм).
studfiles.net
32.Класс Аскомицеты. Общая характеристика. Классификация. Типы плодовых тел. Представители.
Сумчатые грибы, или аскомицеты,— один из крупнейших классов грибов. В нем более 30 000 видов, что составляет около 30% всех известных видов грибов. Входящие в этот класс грибы чрезвычайно разнообразны по строению. Сюда относятся, например, дрожжи, представленные одиночными почкующимися клетками, и виды с плодовыми телами различной формы и размеров, от микроскопических до крупных, достигающих иногда 10—20 см (сморчки, строчки). Но все эти многообразные формы связаны общим происхождением и имеют ряд общих черт, на основании которых они объединяются в этот класс.
Основной признак аскомицетов — образование в результате полового процесса сумок (или асков) — одноклеточных структур, содержащих фиксированное число аскоспор, обычно 8 (рис. 50). Сумки образуются или непосредственно из зиготы (у низших аскомицетов), или на развивающихся из зиготы аскогенных гифах. В сумке происходит слияние ядер зиготы, а затем мейотическое деление диплоидного ядра и образование гаплоидных аскоспор. У высших аскомицетов сумка представляет не только место образования аскоспор, но и активно участвует в их распространении.
Вегетативное тело аскомицетов — разветвленный гаплоидный мицелий, состоящий из многоядерных или одноядерных клеток. В отличие от зигомицетов перегородки (септы) в мицелии аскомицетов образуются упорядоченно, синхронно с делением ядер. Развитие септ происходит от стенок гифы к центру, напоминая сужение диафрагмы в объективе фотоаппарата. В центре септы остается пора (рис. 51), через которую происходит движение цитоплазмы с со скоростью от 1—2 до 25—40 см/ч. Некоторые органеллы клетки, даже ядра, могут мигрировать через поры. Наличие пор в септах играет существенную роль в переносе питательных веществ по гифам в зону роста.
У некоторых аскомицетов мицелий может распадаться на отдельные клетки или почковаться. У дрожжей (порядок Endomycetales) настоящего мицелия нет, а вегетативное тело представлено одиночными почкующимися, реже делящимися клетками, иногда образующими псевдомицелий. Дрожжеподобный рост наблюдается и у некоторых мицелиальных аскомицетов, например у грибов-дерматофитов (порядок Eurotiales), тафриновых(порядок Taphrinales), некоторых видов из рода цератостис (порядок Microascales).
Высокоспециализированные экзопаразиты насекомых из порядка лабульбениевых (Laboulbeniales) имеют редуцированный мицелий, а их тело (рецептакл) состоит из настоящей ткани.
В состав клеточных стенок аскомицетов, как и у хитридиомицетов и зигомицетов, входит хитин, но его содержание у грибов этого класса ниже и составляет не более 20—25% полисахаридов клеточной стенки (для сравнения: у некоторых хитридиомицетов — до 60%, у зигомицетов — до 37—40%). У некоторых дрожжей (род Schizosaccharomyces) хитин не обнаружен. Большую часть полисахаридов клеточной стенки аскомицетов (80—90%) составляют глюканы — полимеры D-глюкозы, отличающиеся от целлюлозы характером связи между мономерами. У дрожжей, кроме глюканов, обнаружены маннаны — полимеры маннозы. Целлюлоза у аскомицетов не обнаружена, за исключением двух видов из рода цератоцистис.
В цикле развития аскомицетов большую роль играет бесполое размножение. Споры бесполого размножения (конидии) образуются на гаплоидном мицелии экзогенно (реже эндогенно) на конидиеносцах различного строения. Конидиальные спороношения аскомицетов очень разнообразны по морфологии. Конидиеносцы образуются одиночно на мицелии, соединяются в пучки (коремии) или подушечки (спородохии), развиваются плотным слоем на поверхности сплетения гиф (ложа) или внутри шаровидных либо грушевидных споровместилищ с отверстием на вершине (пикниды). Типы конидиальных спороношений и различные способы образования конидий подробно онисапы в главе о дейтеромицетах, или несовершенных грибах.
Конидиалыгые спороношения развиваются в период вегетации грибов и служат для их массового расселения. У аскомицетов — паразитов растений они обычно образуются на живом растении, а сумчатые спороношения, за немногими исключениями,— после отмирания растения или его частей, в конце периода вегетации или после перезимовки.
У некоторых аскомицетов конидиальное спороношение неизвестно, у других оно преобладает в цикле развития. В отдельных группах этого класса наблюдается редукция полового процесса и сумчатая стадия образуется редко. Иногда ее трудно обнаружить в природе и получить в искусственной культуре грибов. Поэтому многие аскомицеты как в природе, так и в коллекциях культур мы чаще встречаем в их конидиальной стадии. Это отражается и на их названиях. Конидиальные стадии многих аскомицетов имеют самостоятельные видовые наименования, и в литературе мы чаще встречаем их именно под этими названиями, а не под названиями сумчатых стадий. Например, широко используемый в генетических исследованиях аскомицет Emericella nidulans более известен как Aspergillus nidulans. Распространенный в почве и на различных целлюлозосодержащих субстратах аскомицет Hypocrea rufa обычно называют Trichoderma viride по его конидиальиой стадии. Особенпо распространено это для фитопатогенных аскомицетов: название Fusarium solani вместо Nectria haematococca, Helminthosporium sativum вместо Cochliobolus sativus и т. д. В Международном кодексе ботанической номенклатуры, который регулирует употребление названий растений, содержится даже специальный пункт, разрешающий использовать наряду с основным видовым названием аскомицета (по его сумчатой стадии) название его конидиальной стадии в тех случаях, когда речь идет именно об этой стадии гриба. Это единственный случай, когда для разных стадий в цикле развития одного организма допускаются самостоятельные видовые названия.
Для большого числа грибов, встречающихся в природе в гаплоидной конидиальной стадии, половые стадии неизвестны. Такие грибы относят к классу дейтеромицетов, или несовершенных грибов.
Половой процесс, типичный для аскомицетов,— гаметапгиогамия, т. е. слияние двух гаметангиев — специализированных клеток, не дифференцированных на гаметы.
У низших аскомицетов (подкласс Hemiascomycetidae) половой процесс сходен с зигогамией у зигомицетов. Гаметангии разного пола морфологически сходны или малоразличимы и представляют выросты или веточки мицелия. После их слияния сразу происходит кариогамия и сумка развивается непосредственно из зиготы. Однако, в отличие от зигомицетов, в многоядерных гаметангиях сливаются только два ядра (нет множественной кариогамии), зигота не переходит в состояние покоя, а сразу развивается в сумку. В цикле развития низших аскомицетов, следовательно, есть только гаплоидная и диплоидная стадии.
Для высших аскомицетов (подклассы Euascomycetidae и Loculoascomycetidae) характерны дифференциация и усложнение строения гаметангиев. Образуются одноклеточный антеридий и аскогон, обычно с трихогиной. При оплодотворении содержимое антеридия по трихогине переходит в аскогон. После плазмогамии гаплоидные ядра разного пола не сливаются сразу, а объединяются попарно, образуя дикарионы. Из аскогона вырастают аскогенные гифы, в которых ядра дикариона синхронно делятся. Аскогенные гифы ветвятся и разделяются на двухъядерные клетки. На концах аскогенных гиф развиваются сумки.
Конечная клетка аскогенной гифы загибается крючком, ядра дикариона располагаются в месте перегиба и одновременно делятся. Пара ядер разного пола остается в месте перегиба крючка, одно ядро переходит в его кончик, а другое — в основание. Затем образуются две перегородки, отделяющие одноядерные конечную и базальную клетки крючка. В результате слияния этих клеток восстанавливается дикарион и может происходить повторное образование крючка. Средняя двухъядерная клетка крючка развивается в сумку. Она увеличивается в размерах, ядрадикариона сливаются. Образовавшееся диплоидное ядро делится редукционно, за мейозом следует еще одно, митотическое деление, и вокруг восьми гаплоидных ядер формируются аскоспоры.
По способу образования аскоспоры аскомицетов отличаются от спорангиоспор зародышевого спорангия зигомицетов. Первые образуются по способу так называемого «свободного образования клеток»— часть цитоплазмы сумки обособляется вокруг ядер и одевается оболочкой; при образовании спорангиоспор происходит раскалывание цитоплазмы спорангия. Аскоспоры в сумке окружены не использованной на их формирование цитоплазмой — эпиплазмой. К моменту созревания аскоспор в эпиплазме происходит превращение гликогена в сахар, тургорное давление в сумке возрастает (у некоторых видов — до 10—13 атм, по Ц. Ингольд, 1959) и аскоспоры с силой выбрасываются на расстояние от десятых долей миллиметра до нескольких десятков сантиметров.
Образование аскогенпых гиф увеличивает число сумок, а следовательно, и аскоспор, развивающихся из одного аскогона. Образование сумок по способу крючка с восстановлением дикариона в его базальной клетке и повторным образованием крючков обеспечивает расположение сумок пучком или слоем, что очень важно при активном освобождении аскоспор.
У некоторых аскомицетов (например, из порядка эуроциевых) сумки образуются на аскогенных гифах иными способами — цепочками по ходу аскогенных гиф, из их конечных клеток или боковых выростов.
В цикле развития высших аскомицетов чередуются, таким образом, три стадии: длительная — гаплоидная, в течение которой происходит бесполое размножение, непродолжительная — дикарионтическая (аскогенные гифы) и очень короткая — диплоидная (молодая сумка с диплоидным ядром).
Для многих аскомицетов характерна морфологическая редукция полового процесса. У некоторых представителей этого класса антеридии отсутствуют или не функционируют. В этом случае их функции могут выполнять конидии, вегетативные гифы, а часто мелкие специализированные клетки, называемые спермациями. Спермации нередко образуются на другой особи, на значительном расстоянии от аскогона и переносятся на трихогину токами воздуха, дождем, насекомыми. Трихогина некоторых аскомицетов хемотропична и подрастает к спермациям или конидиям совместимого типа (Ascobolus stercorarius, Podospopa anserina, виды Neurospora). Сперматизация обнаружена в разных группах аскомицетов, например у некоторых гелоциевых, видов из родов Ascobolus и Gelasinospora, в порядке лабульбениевых и у других.
В том случае, когда отсутствуют оба гаметангия, образование дикариона происходит в результате слияния клеток обычных вегетативных гиф одного или двух совместимых мицелиев — соматогамии (например, у Sclerotinia sclerotiorum).
Среди аскомицетов есть гомоталличные и гетероталличные виды, причем гетероталлизм здесь всегда биполярный (двухаллельного типа).
В сумке аскомицетов обычно образуется 8 аскоспор. Однако наблюдаются многочисленные отклонения от тина: например, происходит образование только четырех гаплоидных ядер и соответственно четырех аскоспор (Endomyces magnusii) или часть образовавшихся ядер дегенерирует и число аскоспор за счет этого сокращается (Verpa bohemica). Образование большего числа аскоспор происходит или при большем числе делений ядра (например, у Podospora — семь делений ядра и соответственно 128 аскоспор в сумке), или в результате почкования аскоспор в сумке (например, у тафриновых), или их распада на клетки (у некоторых гипокрейных).
Форма аскоспор очень разнообразна — от шаровидных или эллипсоидальных до нитевидных (Rhytisma, Claviceps). Аскоспоры могут быть одноклеточными или иметь поперечные перегородки, реже — поперечные и продольные перегородки (муральные споры).-
У некоторых аскомицетов аскоспоры имеют разнообразные придатки, играющие роль в их распространении, например слизистые придатки аскоспор копрофильных грибов из рода Podospora или аскоспор сапрофитных морских аскомицетов Ceriosporiopsis halima, Remispora maritima и др.
По строению оболочки и функциям сумки аскомицетов делят на две большие группы — прототуникатные и эутуникатные. Прототуникатные сумки имеют тонкую недифференцированную оболочку, которая разрушается или растворяется, освобождая аскоспоры пассивно. Такая сумка служит только местом формирования спор, но не участвует активно в их распространении. Эутуникатные сумки характеризуются более плотными оболочками, часто со специальными приспособлениями для вскрывания сумки. Они активно участвуют в распространении аскоспор (активное выбрасывание спор из сумки). По строению оболочки эутуникатные сумки могут быть двух типов: унитуникатные и битуникатные. У унитуникатных сумок оболочка относительно тонкая и выглядит однослойной, на вершине сумок имеется обычно апикальный аппарат различного строения, служащий для их вскрывания. Битуникатные сумки имеют ясновыраженную двухслойную оболочку, состоящую из жесткого наружного и эластичного внутреннего слоев. При созревании аскоспор наружный слой оболочки разрушается, начиная с вершины; внутренний слой под действием повышенного тургорного давления растягивается, и происходит активное выбрасывание аскоспор.
Форма сумок округлая или овальная (у гемиаскомицетов и аскомицетов с замкнутыми плодовыми телами) или цилиндрическая (у аскомицетов с активным выбрасыванием аскоспор).
У низших аскомицетов сумки образуются непосредственно на мицелии, а у высших — в специальных вместилищах — плодовых телах (рис. 69). Различают следующие типы плодовых тел: клейстотеций (полностью замкнутое плодовое тело), перитеций (полузамкнутое, обычно кувшиновидпое плодовое тело с отверстием на вершине ) и апотеций (открытое, обычно чашевидное плодовое тело, на верхней стороне которого расположен слой сумок и парафиз). Клейстотеций, перитеции и апотеции — настоящие плодовые тела, развитие которых происходит по аскогимениальному типу — образование их оболочки (перидия) происходит одновременно с развитием аскогенных гиф и сумок.
Настоящие плодовые тела могут развиваться как непосредственно на мицелии, так и на стромах — плотных сплетениях гиф различной формы, размера и консистенции. Однако они всегда имеют собственный перидий, заметный хотя бы на ранних стадиях развития плодового тела.
Кроме настоящих плодовых тел, аскомицеты образуют аскостромы, развитие которых происходит по асколокулярному типу. Сначала закладывается строма из переплетающихся гиф. В ней образуются аскогоны и происходит половой процесс. Аскогенпые гифы и образующиеся на них сумки раздвигают или разрушают плектенхиму стромы, освобождая в ней полость — локулу. Каждая локула содержит одну или несколько сумок. Ткань стромы над локулой разрушается и образуется отверстие, через которое освобождаются аскоспоры. По внешнему виду аскостромы часто похожи на настоящие перитеции, но отличаются от них отсутствием собственного перидия — их оболочкой служит плектенхима стромы.
На основании отсутствия или наличия плодовых тел и способов их образования класс аскомицеты делят на три подкласса:
Подкласс Голосумчатые, или Гемиаскомицеты (Hemiascomycetidae). Плодовые тела отсутствуют. Сумки образуются непосредственно на мицелии, прототуникатные.
Подкласс Эуаскомицеты (Euascomycetidae). Сумки образуются в клейстотециях, перитециях или апотециях, прототуникатные или унитуникатные.
Подкласс Локулоаскомицеты (Loculoascomycetidae). Сумки образуются в аскостромах, битуникатные.
Аскомицеты широко распространены в природе во всех географических областях, на всех доступных для грибов субстратах. Они обитают как сапрофиты в почве, в лесной подстилке, на разнообразных растительных субстратах (древесина, отмершие растения и т. п.). Некоторые группы аскомицетов заняли своеобразные экологические ниши, недоступные для других грибов. Например, кератинофильные грибы, развивающиеся на различных субстратах животного происхождения, содержащих кератин. Некоторые аскомицеты обитают в морях или пресных водоемах на погруженной в воду древесине. Сапрофитные аскомицеты активно участвуют в минерализации органических веществ в природе, особенно в разложении растительных остатков, содержащих целлюлозу. Многие сапрофитные аскомицеты вызывают плесневение и порчу разнообразных материалов и изделий, а также пищевых продуктов.
Многочисленные аскомицеты паразитируют на различных организмах — на растениях (грибах, водорослях, лишайниках и высших), а также на животных и человеке, нередко вызывают серьезные заболевания.
Многие представители этого класса имеют большое экономическое значение как продуценты антибиотиков, алкалоидов, ростовых веществ (гиббереллинов), витаминов (рибофлавина и др.), ферментов, кормового белка, а также как возбудители спиртового брожения. Наконец, многие аскомицеты широко используются сейчас в качестве объектов генетических и биохимических исследований.
studfiles.net
КЛАСС СУМЧАТЫЕ ГРИБЫ ИЛИ АСКОМИЦЕТЫ (ASCOMYCETES)
Сумчатые грибы, или аскомицеты,— один из крупнейших классов грибов. В нем более 30 000 видов, что составляет около 30% всех известных видов грибов. Входящие в этот класс грибы чрезвычайно разнообразны по строению. Сюда относятся, например, дрожжи, представленные одиночными почкующимися клетками, и виды с плодовыми телами различной формы и размеров, от микроскопических до крупных, достигающих иногда 10—20 см (сморчки, строчки). Но все эти многообразные формы связаны общим происхождением и имеют ряд общих черт, на основании которых они объединяются в этот класс.
Основной признак аскомицетов — образование в результате полового процесса сумок (или асков) — одноклеточных структур, содержащих фиксированное число аскоспор, обычно 8 (рис. 50). Сумки образуются или непосредственно из зиготы (у низших аскомицетов), или на развивающихся из зиготы аскогенных гифах. В сумке происходит слияние ядер зиготы, а затем мейотическое деление диплоидного ядра и образование гаплоидных аскоспор. У высших аскомицетов сумка представляет не только место образования аскоспор, но и активно участвует в их распространении.
Вегетативное тело аскомицетов — разветвленный гаплоидный мицелий, состоящий из многоядерных или одноядерных клеток. В отличие от зигомицетов перегородки (септы) в мицелии аскомицетов образуются упорядоченно, синхронно с делением ядер. Развитие септ происходит от стенок гифы к центру, напоминая сужение диафрагмы в объективе фотоаппарата. В центре септы остается пора (рис. 51), через которую происходит движение цитоплазмы с со скоростью от 1—2 до 25—40 см/ч. Некоторые органеллы клетки, даже ядра, могут мигрировать через поры. Наличие пор в септах играет существенную роль в переносе питательных веществ по гифам в зону роста.
У некоторых аскомицетов мицелий может распадаться на отдельные клетки или почковаться. У дрожжей (порядок Endomycetales) настоящего мицелия нет, а вегетативное тело представлено одиночными почкующимися, реже делящимися клетками, иногда образующими псевдомицелий. Дрожжеподобный рост наблюдается и у некоторых мицелиальных аскомицетов, например у грибов-дерматофитов (порядок Eurotiales), тафриновых (порядок Taphrinales), некоторых видов из рода цератостис (порядок Microascales).
Высокоспециализированные экзопаразиты насекомых из порядка лабульбениевых (Laboulbeniales) имеют редуцированный мицелий, а их тело (рецептакл) состоит из настоящей ткани.
В состав клеточных стенок аскомицетов, как и у хитридиомицетов и зигомицетов, входит хитин, но его содержание у грибов этого класса ниже и составляет не более 20—25% полисахаридов клеточной стенки (для сравнения: у некоторых хитридиомицетов — до 60%, у зигомицетов — до 37—40%). У некоторых дрожжей (род Schizosaccharomyces) хитин не обнаружен. Большую часть полисахаридов клеточной стенки аскомицетов (80—90%) составляют глюканы — полимеры D-глюкозы, отличающиеся от целлюлозы характером связи между мономерами. У дрожжей, кроме глюканов, обнаружены маннаны — полимеры маннозы. Целлюлоза у аскомицетов не обнаружена, за исключением двух видов из рода цератоцистис.
В цикле развития аскомицетов большую роль играет бесполое размножение. Споры бесполого размножения (конидии) образуются на гаплоидном мицелии экзогенно (реже эндогенно) на конидиеносцах различного строения. Конидиальные спороношения аскомицетов очень разнообразны по морфологии. Конидиеносцы образуются одиночно на мицелии, соединяются в пучки (коремии) или подушечки (спородохии), развиваются плотным слоем на поверхности сплетения гиф (ложа) или внутри шаровидных либо грушевидных споровместилищ с отверстием на вершине (пикниды). Типы конидиальных спороношений и различные способы образования конидий подробно онисапы в главе о дейтеромицетах, или несовершенных грибах.
Конидиалыгые спороношения развиваются в период вегетации грибов и служат для их массового расселения. У аскомицетов — паразитов растений они обычно образуются на живом растении, а сумчатые спороношения, за немногими исключениями,— после отмирания растения или его частей, в конце периода вегетации или после перезимовки.
У некоторых аскомицетов конидиальное спороношение неизвестно, у других оно преобладает в цикле развития. В отдельных группах этого класса наблюдается редукция полового процесса и сумчатая стадия образуется редко. Иногда ее трудно обнаружить в природе и получить в искусственной культуре грибов. Поэтому многие аскомицеты как в природе, так и в коллекциях культур мы чаще встречаем в их конидиальной стадии. Это отражается и на их названиях. Конидиальные стадии многих аскомицетов имеют самостоятельные видовые наименования, и в литературе мы чаще встречаем их именно под этими названиями, а не под названиями сумчатых стадий. Например, широко используемый в генетических исследованиях аскомицет Emericella nidulans более известен как Aspergillus nidulans. Распространенный в почве и на различных целлюлозосодержащих субстратах аскомицет Hypocrea rufa обычно называют Trichoderma viride по его конидиальиой стадии. Особенпо распространено это для фитопатогенных аскомицетов: название Fusarium solani вместо Nectria haematococca, Helminthosporium sativum вместо Cochliobolus sativus и т. д. В Международном кодексе ботанической номенклатуры, который регулирует употребление названий растений, содержится даже специальный пункт, разрешающий использовать наряду с основным видовым названием аскомицета (по его сумчатой стадии) название его конидиальной стадии в тех случаях, когда речь идет именно об этой стадии гриба. Это единственный случай, когда для разных стадий в цикле развития одного организма допускаются самостоятельные видовые названия.
Для большого числа грибов, встречающихся в природе в гаплоидной конидиальной стадии, половые стадии неизвестны. Такие грибы относят к классу дейтеромицетов, или несовершенных грибов.
Половой процесс, типичный для аскомицетов,— гаметапгиогамия, т. е. слияние двух гаметангиев — специализированных клеток, не дифференцированных на гаметы.
У низших аскомицетов (подкласс Hemiascomycetidae) половой процесс сходен с зигогамией у зигомицетов. Гаметангии разного пола морфологически сходны или малоразличимы и представляют выросты или веточки мицелия. После их слияния сразу происходит кариогамия и сумка развивается непосредственно из зиготы. Однако, в отличие от зигомицетов, в многоядерных гаметангиях сливаются только два ядра (нет множественной кариогамии), зигота не переходит в состояние покоя, а сразу развивается в сумку. В цикле развития низших аскомицетов, следовательно, есть только гаплоидная и диплоидная стадии (рис. 52).
Для высших аскомицетов (подклассы Euascomycetidae и Loculoascomycetidae) характерны дифференциация и усложнение строения гаметангиев. Образуются одноклеточный антеридий и аскогон, обычно с трихогиной. При оплодотворении содержимое антеридия по трихогине переходит в аскогон. После плазмогамии гаплоидные ядра разного пола не сливаются сразу, а объединяются попарно, образуя дикарионы. Из аскогона вырастают аскогенные гифы, в которых ядра дикариона синхронно делятся. Аскогенные гифы ветвятся и разделяются на двухъядерные клетки. На концах аскогенных гиф развиваются сумки (см. рис. 50).
Конечная клетка аскогенной гифы загибается крючком, ядра дикариона располагаются в месте перегиба и одновременно делятся. Пара ядер разного пола остается в месте перегиба крючка, одно ядро переходит в его кончик, а другое — в основание. Затем образуются две перегородки, отделяющие одноядерные конечную и базальную клетки крючка. В результате слияния этих клеток восстанавливается дикарион и может происходить повторное образование крючка. Средняя двухъядерная клетка крючка развивается в сумку. Она увеличивается в размерах, ядрадикариона сливаются. Образовавшееся диплоидное ядро делится редукционно, за мейозом следует еще одно, митотическое деление, и вокруг восьми гаплоидных ядер формируются аскоспоры.
По способу образования аскоспоры аскомицетов отличаются от спорангиоспор зародышевого спорангия зигомицетов. Первые образуются по способу так называемого «свободного образования клеток»— часть цитоплазмы сумки обособляется вокруг ядер и одевается оболочкой; при образовании спорангиоспор происходит раскалывание цитоплазмы спорангия. Аскоспоры в сумке окружены не использованной на их формирование цитоплазмой — эпиплазмой. К моменту созревания аскоспор в эпиплазме происходит превращение гликогена в сахар, тургорное давление в сумке возрастает (у некоторых видов — до 10—13 атм, по Ц. Ингольд, 1959) и аскоспоры с силой выбрасываются на расстояние от десятых долей миллиметра до нескольких десятков сантиметров.
Образование аскогенпых гиф увеличивает число сумок, а следовательно, и аскоспор, развивающихся из одного аскогона. Образование сумок по способу крючка с восстановлением дикариона в его базальной клетке и повторным образованием крючков обеспечивает расположение сумок пучком или слоем, что очень важно при активном освобождении аскоспор.
У некоторых аскомицетов (например, из порядка эуроциевых) сумки образуются на аскогенных гифах иными способами — цепочками по ходу аскогенных гиф, из их конечных клеток или боковых выростов.
В цикле развития высших аскомицетов чередуются, таким образом, три стадии: длительная — гаплоидная, в течение которой происходит бесполое размножение, непродолжительная — дикарионтическая (аскогенные гифы) и очень короткая — диплоидная (молодая сумка с диплоидным ядром).
Для многих аскомицетов характерна морфологическая редукция полового процесса. У некоторых представителей этого класса антеридии отсутствуют или не функционируют. В этом случае их функции могут выполнять конидии, вегетативные гифы, а часто мелкие специализированные клетки, называемые спермациями. Спермации нередко образуются на другой особи, на значительном расстоянии от аскогона и переносятся на трихогину токами воздуха, дождем, насекомыми. Трихогина некоторых аскомицетов хемотропична и подрастает к спермациям или конидиям совместимого типа (Ascobolus stercorarius, Podospopa anserina, виды Neurospora). Сперматизация обнаружена в разных группах аскомицетов, например у некоторых гелоциевых, видов из родов Ascobolus и Gelasinospora, в порядке лабульбениевых и у других.
В том случае, когда отсутствуют оба гаметангия, образование дикариона происходит в результате слияния клеток обычных вегетативных гиф одного или двух совместимых мицелиев — соматогамии (например, у Sclerotinia sclerotiorum).
Среди аскомицетов есть гомоталличные и гетероталличные виды, причем гетероталлизм здесь всегда биполярный (двухаллельного типа).
В сумке аскомицетов обычно образуется 8 аскоспор. Однако наблюдаются многочисленные отклонения от тина: например, происходит образование только четырех гаплоидных ядер и соответственно четырех аскоспор (Endomyces magnusii) или часть образовавшихся ядер дегенерирует и число аскоспор за счет этого сокращается (Verpa bohemica). Образование большего числа аскоспор происходит или при большем числе делений ядра (например, у Podospora — семь делений ядра и соответственно 128 аскоспор в сумке), или в результате почкования аскоспор в сумке (например, у тафриновых), или их распада на клетки (у некоторых гипокрейных).
Форма аскоспор очень разнообразна — от шаровидных или эллипсоидальных до нитевидных (Rhytisma, Claviceps). Аскоспоры могут быть одноклеточными или иметь поперечные перегородки, реже — поперечные и продольные перегородки (муральные споры).-
У некоторых аскомицетов аскоспоры имеют разнообразные придатки, играющие роль в их распространении, например слизистые придатки аскоспор копрофильных грибов из рода Podospora или аскоспор сапрофитных морских аскомицетов Ceriosporiopsis halima, Remispora maritima и др.
По строению оболочки и функциям сумки аскомицетов делят на две большие группы — прототуникатные и эутуникатные. Прототуникатные сумки имеют тонкую недифференцированную оболочку, которая разрушается или растворяется, освобождая аскоспоры пассивно. Такая сумка служит только местом формирования спор, но не участвует активно в их распространении. Эутуникатные сумки характеризуются более плотными оболочками, часто со специальными приспособлениями для вскрывания сумки. Они активно участвуют в распространении аскоспор (активное выбрасывание спор из сумки). По строению оболочки эутуникатные сумки могут быть двух типов: унитуникатные и битуникатные. У унитуникатных сумок оболочка относительно тонкая и выглядит однослойной, на вершине сумок имеется обычно апикальный аппарат различного строения, служащий для их вскрывания. Битуникатные сумки имеют ясновыраженную двухслойную оболочку, состоящую из жесткого наружного и эластичного внутреннего слоев. При созревании аскоспор наружный слой оболочки разрушается, начиная с вершины; внутренний слой под действием повышенного тургорного давления растягивается, и происходит активное выбрасывание аскоспор.
Форма сумок округлая или овальная (у гемиаскомицетов и аскомицетов с замкнутыми плодовыми телами) или цилиндрическая (у аскомицетов с активным выбрасыванием аскоспор).
У низших аскомицетов сумки образуются непосредственно на мицелии, а у высших — в специальных вместилищах — плодовых телах (рис. 69). Различают следующие типы плодовых тел: клейстотеций (полностью замкнутое плодовое тело), перитеций (полузамкнутое, обычно кувшиновидпое плодовое тело с отверстием на вершине ) и апотеций (открытое, обычно чашевидное плодовое тело, на верхней стороне которого расположен слой сумок и парафиз). Клейстотеций, перитеции и апотеции — настоящие плодовые тела, развитие которых происходит по аскогимениальному типу — образование их оболочки (перидия) происходит одновременно с развитием аскогенных гиф и сумок.
Настоящие плодовые тела могут развиваться как непосредственно на мицелии, так и на стромах — плотных сплетениях гиф различной формы, размера и консистенции. Однако они всегда имеют собственный перидий, заметный хотя бы на ранних стадиях развития плодового тела.
Кроме настоящих плодовых тел, аскомицеты образуют аскостромы, развитие которых происходит по асколокулярному типу. Сначала закладывается строма из переплетающихся гиф. В ней образуются аскогоны и происходит половой процесс. Аскогенпые гифы и образующиеся на них сумки раздвигают или разрушают плектенхиму стромы, освобождая в ней полость — локулу. Каждая локула содержит одну или несколько сумок. Ткань стромы над локулой разрушается и образуется отверстие, через которое освобождаются аскоспоры. По внешнему виду аскостромы часто похожи на настоящие перитеции, но отличаются от них отсутствием собственного перидия — их оболочкой служит плектенхима стромы.
На основании отсутствия или наличия плодовых тел и способов их образования класс аскомицеты делят на три подкласса:
Подкласс Голосумчатые, или Гемиаскомицеты (Hemiascomycetidae). Плодовые тела отсутствуют. Сумки образуются непосредственно на мицелии, прототуникатные.
Подкласс Эуаскомицеты (Euascomycetidae). Сумки образуются в клейстотециях, перитециях или апотециях, прототуникатные или унитуникатные.
Подкласс Локулоаскомицеты (Loculoascomycetidae). Сумки образуются в аскостромах, битуникатные.
Аскомицеты широко распространены в природе во всех географических областях, на всех доступных для грибов субстратах. Они обитают как сапрофиты в почве, в лесной подстилке, на разнообразных растительных субстратах (древесина, отмершие растения и т. п.). Некоторые группы аскомицетов заняли своеобразные экологические ниши, недоступные для других грибов. Например, кератинофильные грибы, развивающиеся на различных субстратах животного происхождения, содержащих кератин. Некоторые аскомицеты обитают в морях или пресных водоемах на погруженной в воду древесине. Сапрофитные аскомицеты активно участвуют в минерализации органических веществ в природе, особенно в разложении растительных остатков, содержащих целлюлозу. Многие сапрофитные аскомицеты вызывают плесневение и порчу разнообразных материалов и изделий, а также пищевых продуктов.
Многочисленные аскомицеты паразитируют на различных организмах — на растениях (грибах, водорослях, лишайниках и высших), а также на животных и человеке, нередко вызывают серьезные заболевания.
Многие представители этого класса имеют большое экономическое значение как продуценты антибиотиков, алкалоидов, ростовых веществ (гиббереллинов), витаминов (рибофлавина и др.), ферментов, кормового белка, а также как возбудители спиртового брожения. Наконец, многие аскомицеты широко используются сейчас в качестве объектов генетических и биохимических исследований.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
dic.academic.ru
Аскомицетовые дрожжи
К аскомицетам относят дрожжи, половые споры которых формируются эндогенно внутри особых вместилищ - сумок, или асков. В отличие от мицелиальных аскомицетов, большая часть жизненного цикла которых проходит в гаплоидной фазе, а диплоидна только молодая сумка, среди аскоспоровых дрожжей есть виды с разными типами онтогенеза: гаплоидным, диплоидным и гапло-диплоидным. При этом аском может стать либо непосредственно зигота (Schizosaccharomyce, Zygosaccharomyces), либо отдельная диплоидная вегетативная клетка (Saccharomyces, Saccharomycodes), либо сумка развивается как новообразование с участием так называемых «активных» почек, выполняющих функцию гамет (Lipomyces).
Аски могут также формироваться из хламидоспор, из клеток псевдомицелия, или на истинном мицелии. В любом случае формирование аска происходит из диплоидной клетки после мейотического деления ядра. Мейоз представляет собой два следующих друг за другом деления ядра с однократным удвоением хромосом. Перед вторым делением хромосомы не удваиваются, поэтому в результате происходит редукция числа хромосом и возникают четыре гаплоидных ядра. Затем в районе ядерных бляшек начинают формироваться мембраны, которые постепенно разрастаются и охватывают часть цитоплазмы будущего аска. После образования аскоспор остается небольшое количество «неиспользованной» цитоплазмы. Между двумя листками мембран закладывается клеточная стенка аскоспоры. Аскоспоры отличаются от вегетативных клеток более толстой и многослойной клеточной стенкой, меньшим развитием внутриклеточных мембранных систем, отсутствием вакуолей. Эти особенности связаны с пониженной метаболической активностью аскоспор.
 Обычно аскоспоры содержат большое количество запасных веществ, чаще всего - липидов. Аскоспоры дрожжей - это типичные покоящиеся споры, способные более или менее длительный период существовать в неактивном состоянии.
Обычно аскоспоры содержат большое количество запасных веществ, чаще всего - липидов. Аскоспоры дрожжей - это типичные покоящиеся споры, способные более или менее длительный период существовать в неактивном состоянии. Однако, устойчивость аскоспор к ряду повреждающих факторов, например, к повышенным температурам, обычно не намного выше, чем у вегетативных клеток. Форма аскоспор у дрожжей очень разнообразна. Они могут быть круглыми, овальными, бобовидными, чечевицеобразными, серповидными, игловидными и т.д. Кроме того, аскоспоры могут иметь на поверхности клеточной стенки различные скульптурные образования, которые хорошо выявляются в электронном микроскопе. За счет таких образований аскоспоры могут быть бородавчатыми, сатурновидными, напоминающими грецкий орех и пр.
Морфология асков зависит от способа их образования, типа полового процесса (гологамия или педогамия), количества и формы аскоспор. Количество аскоспор в аске у разных видов может быть от одной до нескольких десятков. Наиболее часто встречаются аски с 1,2,4 и 8 аскоспорами. Аски различаются также временем существования. У большинства дрожжей аски устойчивые и разрушаются только в очень старых культурах. Но есть виды с быстро разрушающимися асками. Морфология асков, аскоспор, тип полового процесса имеют большое таксономическое значение. Эти признаки используются в систематике дрожжей при выделении таксонов родового уровня. Рассмотрим несколько примеров жизненных циклов аскомицетовых дрожжей.
· Гаплоидные. У таких дрожжей вегетативное размножение происходит в гаплоидной фазе, а диплоидная стадия очень короткая: образовавшееся диплоидное ядро сразу же делится мейотически с восстановлением гаплоидного состояния. Schizosaccharomyces pombe. Половой процесс - гологамия. Две морфологически сходные гаплоидные вегетативные клетки образуют выросты, с помощью которых происходит контакт, а затем слияние содержимого клеток. Возникает диплоидная зигота, которая вегетативно не размножается, а ядро ее сразу переходит к мейотическому делению. Образующиеся четыре гаплоидных ядра включаются в аскоспоры. После освобождения из аска аскоспоры прорастают и дают начало длительной стабильной вегетативной фазе. Lipomyces tetrasporus. Половой процесс - адельфогамия. Роль гамет здесь выполняют активные почки. Такие почки формируются на гаплоидных вегетативных клетках на поздних стадиях роста после периода вегетативного размножения. Обычно две почки на одной материнской клетке выполняют функцию гамет и копулируют между собой, образуя зиготу, которая затем разрастается в виде мешка и отделяется перегородкой от несущей ее клетки. Ядро зиготы делится мейотически и она превращается в четырехспоровый мешковидный аск, прикрепленный к материнской клетке, на которой может затем формироваться вторая и третья сумки.
· Диплоидные. У этих дрожжей вегетативно размножаются только диплоидные клетки. Гаплофаза ограничена молодыми асками и аскоспорами. . Saccharomycodes ludwigii. У этих дрожжей диплоидизация происходит при слиянии аскоспор. Четыре гаплоидные аскоспоры прорастают и начинают копулировать попарно, когда они еще находятся в аске. Образовавшиеся диплоидные клетки размножаются вегетативно, образуя стабильную и длительную диплофазу. При соответствующих условиях, когда снимается контроль митотического деления ядра, диплоидная клетка вступает в митотический цикл и превращается в аск с 4 аскоспорами. Hanseniaspora uvarum. Жизненный цикл сходен с описанным выше за исключением того, что споры не копулируют, но ядро в зрелой споре после освобождения ее из сумки в условиях, обеспечивающих вегетацию, делится мейотически. Образовавшиеся два гаплоидных ядра сливаются, образуя уже диплоидную клетку, способную к вегетативному размножению. Сходный цикл наблюдается и у почвенных дрожжей Williopsis saturnus.
· Гапло-диплоидные. Существуют дрожжи, у которых вегетативное размножение может происходить как в гаплоидной, так и в диплоидной фазах. Длительность той или другой фазы зависит от вида и от условий роста. Saccharomyces cerevisiae. Эти дрожжи вегетируют преимущественно в диплоидном состоянии, но у них имеется короткая вегетативная гаплоидная фаза. Диплоидная клетка в условиях дефицита легкодоступных источников углерода прекращает почковаться, и ядро ее делится мейотически. В результате она превращается в аск с 4 гаплоидными аскоспорами, которые после освобождения из аска прорастают и образуют гаплоидное поколение. Гаплоидные клетки обычно мельче диплоидных и имеют более округлую форму. Шрамы почкования у них сближены, почки образуются группами в одном локусе. После нескольких циклов почкования две клетки конъюгируют и сливаются, восстанавливая диплоидное состояние.
biofile.ru
Настоящие грибы. Отдел Аскомицеты – Ascomycota
Аскомицеты составляют до 30% всех видов грибов – около 30000. К ним можно отнести еще и дейтеромицеты, у котороых был обнаружен половой процесс, и сумчатые лишайники – тогда число видов достигает почти половины из всех известных. Экологические группы очень разнообразны.
Основное таксономическое отличие аскомицетов – формирование сумки, или аска, в которой происходит кариогамия, мейоз и дифференцировка мейоспор – аскоспор.
Основные полисахариды клеточной стенки – хитин и глюканы. Хитин иногда вовсе отсутствует (дрожжи). Таллом представлен ветвящимся гаплоидным мицелием из одноядерных или многоядерных клеток. Септы простые, образуются центрипетально синхронно с делением клеток. У низших аскомицетов (тафриномицеты, гемиаскомицеты) вегетативный таллом может быть представлен одной клеткой, типичного мицелия часто нет, развивается псевдомицелий. И, наоборот, иногда и высшие аскомицеты (эуаскомицеты) могут развивать дрожжеподобный мицелий (мицелиально-дрожжевой диморфизм).
Вегететативное размножение: фрагментами таллома.
Бесполое размножение: конидиальные спороношения разного типа (одиночные, коремии, спородохии и др.). Развиваются в период вегетации грибов и служат для расселения вида.
Аскомицеты, у которых в жизненном цикле преобладает бесполое, конидиальное размножение, а половое встречалось в единичных случаях или вовсе неизвестно, входят сразу в две таксономические группы: в группу аскомицетов и в группу несовершенных грибов – дейтеромицетов, причем часто даже под разными названиями. Так, один из аскомицетов в стадии полового спороношения (телеоморфа) носит название Emericella nidulans, а в стадии конидиального спороношения (анаморфа) – Aspergillus nidulans.
Тип полового процесса – чаще всего гаметангиогамия, реже соматогамия или сперматизация. Есть виды и гомо-, и гетероталличные. В цикле развития аскомицетов чередуется не две, а три ядерных фазы: длительная и основная – гаплоидная (вегетативный мицелий), непродолжительная дикариотическая (аскогенные гифы) и очень короткая диплоидная (сумка с диплоидным ядром).
У низших аскомицетов половой процесс сходен с зигогамией: разнополые гаметангии морфологически сходны. Сразу после слияния происходит кариогамия, мейоз и развитие сумки – дикариотической фазы нет. У высших гаметангии четко дифференцированы. Женсий гаметангий состоит из округлого аскогона и удлиненной трихогины, мужской – антеридий – одноклеточный. При слиянии содержимое антеридия по трихогине переходит в аскогон. В нем происходит плазмогамия и образуется дикарион – объединение генетически разнородных ядер в пару пока без окончательного слияния. Из аскогона вырастают аскогенные гифы, каждая из которых формируют от одной до множества сумок, или асков (двумя способами: крючком или цепочкой).
В сумке ядра диканриона сливаются – происходит кариогамия, и мейоз с образованием 4 ядер, которые затем делятся митотически. В итоге образуется 8 гаплоидных ядер, вокруг каждого обособляется часть цитоплазмы сумки и формируются оболочки; так ядра становятся аскоспорами. В сумке они окружены неиспользованной на их образование цитоплазмой – эпиплазмой. Ко времени созревания аскоспор гликоген в ней переходит в сахар, тургорное давление в сумкке возрастает и аскоспоры резко выбрасываются на расстояние до 10 см. Аскоспоры могут очень различаться по форме, размерам и числу клеток, в сумке их может быть от 1 до 128, чаще 8.
Выделяется несколько типов сумок:
1. Прототуникатные – с тонкой недифференцированной оболочкой. Они разрушаются, пассивно высвобождая споры.
2. Эутуникатные – имеют более плотные оболочки с приспособлением для вскрывания сумки, активно участвуют в распространении спор. Строение оболочки двух типов:
- унитуникатные сумки – оболочка однослойная тонкая с апикальным аппаратом различного строения на вершине
- битуникатные сумки – оболочка двуслойная: жесткий наружный и эластичный внутренний. При созревании спор сначала с вершины разрушается наружный слой, а затем от повышающегося тургорного давления растягивается и разрывается внутрений, активно выбрасывая аскоспоры.
У низших аскомицетов сумки образуются прямо на мицелии, у высших – в специальных образованиях – плодовых телах и аскостромах.
Настоящие плодовые тела образуются после плазмогамии и полового процесса и всегда имеют собственный перидий (оболочку): гаплоидные вегетативные гифы оплетают аскогенные гифы и сумки, образуя довольно плотные структуры. Сумки в гимениальном слое плодовых тел перемежаются стерильными элементами – парафизами. Они отделяют сумки друг от друга и способствуют распространению спор, предохраняют их от слипания.
Плодовые тела различаются по степени замкнутости:
1. Клейстотеции – полностью замкнутое округло-шаровидное плодовое тело.
2. Перитеций – полузамкнутое, обычно кувшиновидное плодовое тело с отверстием на вершине.
3. Апотеций – открытое чашевидное плодовое тело.
Образование стромы характерно для многих групп аскомицетов. Она представляет собой компактное образование из плотно сплетенных гиф, в котором или на котором располагаются несущие споры структуры полового, бесполого или обоих типов размножения. В отличие от склероция строма, как правило, не является покоящейся структурой. В другом отделе высших грибов – базидиомицеты – стромы образуются крайне редко, лишь как исключение.
У аскомицетов строма особенно часто встречается в классе сордариомицеты (ранее пиреномицеты). В этом классе наблюдаются настоящие плодовые тела – перитеции, погруженные в строму или расположенные на ней.
У дотидеомицетов (ранее локулоаскомицеты) образуются аскостромы, которые, как правило, считаются ложными плодовыми телами. Аскострома формируется перед плазмогамией и половым процессом. Т.е. сначала гифы переплетаются и образуют строму, затем в ней образуются аскогоны и антеридии и происходит половой процесс. Затем появившиеся сумки расширяют паренхиму стромы, создавая в ней полости – локулы. Над локулой строма разрушается, создавая отверстие. Внешне аскостромы могут быть похожи на настоящие перитеции. Такие аскостромы называют псевдотециями.
Система аскомицетов, как, впрочем, и всех остальных грибов, в настоящее время претерпевает существенные изменения. Меняется число и объем порядков, подклассов и даже классов. Классификация основана на типах сумок и плодовых тел. Выделяется 3 подотдела и около 10 классов:
Подотдел тафриномицеты – Taphrinomycotina – мицелиальные, почкующиеся или диморфные формы. Сумки прототуникатные, расположены слоем. Плодовые тела образуются крайне редко.
Подотдел гемиаскомицеты – Hemiascomycotina – как правило, почкующиеся, реже диморфные и мицелиальные формы. Сумки или прототуникатные, или развиваются как одиночные свободные клетки. Плодовых тел нет.
Подотдел эуаскомицеты – Ascomycotina – мицелиальные формы с настоящими или ложными плодовыми телами.
1. Класс эвроциомицеты – Eurotiomycetes – плодовые тела – клейстотеции с беспорядочно расположенными прототуникатными сумками.
2. Класс эризифомицеты – Erysiphomycetes – плодовые тела – клейстотеции с упорядоченно расположенными унитуникатными сумками.
3. Класс сордариомицеты – Sordariomycetes – плодовые тела – перитеции с расположенными пучком или слоем унитуникатными сумками.
4. Класс пезизомицеты – Pezizomycetes – плодовые тела – апотеции с расположенным слоем унитуникатными оперкулятными сумками
5. Класс леоциомицеты – Leotiomycetes – плодовые тела – апотеции с расположенными слоем унитуникатными иноперкулятными сумками
6. Класс леканоромицеты – Lecanoromycetales – лихенизированные грибы. Плодовые тела – апотеции с расположенными слоем леканоровыми сумками (модифицированные битуникатные).
7. Класс дотидеомицеты – Dothideomycetes – битуникатные сумки расположены в локулах.
biofile.ru
Аскомицеты | Virtual Laboratory Wiki
Аскомицеты (от греч. ἀσκός — сумка), сумчатые грибы (лат. Ascomycota) — отдел в царстве грибов, объединяющий организмы с септированным мицелием и специфическими органами полового спороношения — сумками (асками), содержащими чаще всего по 8 аскоспор. Имеют и бесполое спороношение, причём во многих случаях половой процесс утрачивается и такие виды грибов относят к несовершенным грибам (Deuteromycota).
К аскомицетам относят до 2000 родов и 30 000 видов. Среди них — дрожжи (класс Saccharomycetes) — вторично одноклеточные организмы. Из других известных представителей аскомицетов можно назвать сморчки, строчки и трюфели.
Мицелий разделён поперечными перегородками (септирован), что позволяет гифе в случае повреждения терять меньше клеточного содержимого и обуславливает большую выживаемость аскомицетов по сравнению с ценоцитными зигомицетами. Так, наименьший жизнеспособный фрагмент мицелия, способный дать начало новому организму, для зигомицета составляет около 100 мкм, а для аскомицета — 15-20 мкм (1-3 целых клетки). В середине септы имеется простая пора, через которую возможен обмен цитоплазмой, органеллами и даже ядрами. Чаще всего у аскомицетов одно гаплоидное ядро в клетке.
Размножение и цикл развития Править
У аскомицетов выражены две стадии развития: анаморфа, в которой организм размножается бесполым путём, и телиоморфа, в которой он формирует половые структуры. Вместе обе стадии носят название голоморфа. Анаморфа и телиоморфа не схожи, в зависимости от условий организм может переходить в телиоморфу, а может и нет, в результате чего часто один и тот же аскомицет описывался как два разных вида. Например, название Aspergillus fisheri дано анаморфе, телиоморфа которой описана как Neosartoria fisheri, аналогичная ситуация с Amorphotheca resinae и Hormoconis resinae и многими другими. В наши дни молекулярные методы позволяют обнаружить описанные дважды виды.
Бесполое размножение Править
Бесполое размножение аскомицетов осуществляется конидиями, которые образуются экзогенно на конидиеносцах, собранных в группы. Скопления конидиеносцев могут представлять собой пучки (коремии), подушечки (спородохии), либо образовывать слой на сплетении гиф — ложе.
Половое размножение Править
Половое размножение аскомицетов проходит в форме гаметангиогамии, без образования дифферинцированных гамет. Для этого на разных гифах образуются половые органы: мужские — антеридии и женские — архикарпы, представляющие собой видоизменённые оогонии.
У низших форм половой процесс напоминает зигогамию у зигомицетов (Zygomycota), поскольку образующиеся гаметангии внешне сходны и после их объединения сразу происходит слияние ядер. Однако у низших асковых грибов сливаются только два ядра многоядерных гаметангиев, а не все, то есть отсутствует множественная кариогамия. Образовавшееся диплоидное ядро без периода покоя делится мейотически, образуя гаплоидные ядра, а зигота трансформируется в сумку — аск (греч. askos — мешок).
У высших форм половой процесс проходит более сложно. Архикарп у них дифференцирован на расширенную нижнюю часть — аскогон и верхнюю в виде изогнутой трубочки — трихогину. Антеридий представляет собой одноклеточную структуру цилиндрической формы. При соприкосновении гаметангиев трихогина врастает своим концом в антеридий, после чего содержимое антеридия перетекает по ней в аскогон. Слияние цитоплазмы гаметангиев (плазмогамия) не сопровождается слиянием их гаплоидных ядер, хотя они сближаются и располагаются попарно, образуя дикарионы. Затем из аскогона вырастают аскогенные гифы, одновременно ядра дикарионов делятся, что ведёт к увеличению числа дикарионов. В каждую из аскогенных гиф проникают дикарионы. Завершается процесс образованием сумок (асков), которыми становятся концевые клетки, расположенные на концах аскогенных гиф, содержащие дикарион. Вначале гаплоидные ядра дикариона сливаются (происходит кариогамия), образуя диплоидное ядро. Без периода покоя это ядро делится мейотически, образуя четыре гаплоидных ядра, а те, в свою очередь, делятся митотически. В итоге появляются восемь гаплоидных клеток, которые становятся аскоспорами, а клетка, в которой они находятся, становится аском. У некоторых видов количество спор может быть меньшим (четыре) за счёт отсутствия митотического деления после мейотического или за счёт дегенерации части гаплоидных ядер или большим.
Таким образом, в жизненном цикле асковых грибов имеется три стадии: гаплоидная стадия, когда мицелий размножается бесполым путём; стадия дикариона и самая короткая — диплоидная стадия, когда молодая сумка непродолжительное время содержит диплоидное ядро.
У многих сумчатых грибов половой процесс упрощается. У них не образуются антеридии, а вместо них функционируют вегетативные гифы, конидии или мелкие клетки спермации, которые могут образовываться даже не на соседних гифах, а на достаточном расстоянии от архикарпа. Потоками воздуха, воды или через насекомых спермации переносятся на трихогину, после чего происходит слияние цитоплазмы клеток. Случается, что могут отсутствовать оба гаметангия, тогда половой процесс протекает в форме соматогамии, то есть сливаются вегетативные клетки. При этом следует отметить, что у сумчатых грибов есть формы с гомоталличным и гетероталличным мицелиями, причём последние всегда отличаются набором аллелей.
Аскогенные гифы с асками могут образовываться не только беспорядочно (в любом месте мицелия), как это происходит у низших форм, но и на плодовых телах, состоящих из плотно переплетённых гиф. У сумчатых грибов имеются три типа плодовых тел: <span />
- Клейстотеции представляют собой полностью замкнутое плодовое тело с находящимися внутри асками, освобождающимися после разрушения его стенок. Характерны для группы порядков плектомицеты.
- Перитеции (греч. peri — возле, около, theke — сумка) — обычно имеют кувшинообразную форму с отверстием в верхней части. Характерны для группы порядков пиреномицеты.
- Апотеции — открытые вместилища аск. Образуют чаши или сморщенную поверхность сморчков (причём рёбра складок стерильны), могут, однако, не раскрываться (у трюфелей). Аскомицеты с апотециями считаются наиболее высокоорганизованными и объединяются в группу порядков дискомицеты.
Клейстотеции и перитеции часто развиваются на особых образованиях — стромах, которые часто ошибочно принимают за плодовые тела. Они могут находиться на поверхности стромы или быть погружёнными в её тело.
Выделяют 4 класса аскомицетов:
К истинным аскомицетам относятся более 90 % видов из отдела Ascomycota. По строению плодовых тел и сумок их разделяют на 40 порядков.
Значение в природе и жизни человека Править
Аскомицеты играют важную роль в наземных экосистемах. Разрушая такие субстраты, как опад, мёртвые ветки и стволы деревьев (в том числе входящие в их состав целлюлозу и лигнин, недоступные большинству организмов), они вносят существенный вклад в биологические циклы углерода и азота. Сами по себе аскомицеты являются пищей для беспозвоночных, грызунов и более крупных животных: оленей и кабанов.
Аскомицеты являются основой для многих (до 98 %) лишайников. С корневыми системами высших растений многие аскомицеты образуют микоризу. Грибы рода Xylaria культивируются муравьями Attini и некоторыми термитами.
Многие асковые широко используются в хозяйственной деятельности человека, например, трюфели и сморчки употребляются в пищу. Дрожжи незаменимы в хлебопечении и на бродильных производствах, другие виды широко используются для получения биологически активных веществ (антибиотиков, витаминов, ферментов, алкалоидов, гиббереллинов и пр.). Многие виды широко используются в генетических исследованиях.
- Сидорова И.И. и др. Класс Аскомицеты (Ascomycetes) // Мир растений. в 7 т. / Редкол. А.Л. Тахтаджян (гл. ред.) и др.. Т. 2. Грибы. / Под ред. М.В. Горленко. 2-е изд., перераб. - М.: Просвещение, 1991. - 475 с., 24 л.: ил. - ISBN 5-09-002851-9
Эта страница использует содержимое раздела Википедии на русском языке. Оригинальная статья находится по адресу: Аскомицеты. Список первоначальных авторов статьи можно посмотреть в истории правок. Эта статья так же, как и статья, размещённая в Википедии, доступна на условиях CC-BY-SA .
ru.vlab.wikia.com
Класс Аскомицеты, или сумчатые грибы Ascomycetes
Аскомицеты, или сумчатые грибы – один из обширнейших классов грибов, включающих 30 000 видов, разнообразных по строению, та и по образу жизни. Основные признаки аскомицетов формирование в результате полового процесса сумок, или асков, — замкнутых одноклеточных структур, содержащих определенное количество аскоспор, обычно восемь.
Вегетативное тело аскомицетов – разветвленный гаплоидный мицелий, состоящий из одноядерных или многоядерных клеток. Септы образуются упорядоченно, синхронно с делением ядер. Септа развивается от стенок гифа, к центру и напоминает сужение ирисовой диафрагмы в объективе фотоаппарата. В центре септы остается пора, через которую передвигается цитоплазма, а так же могут мигрировать органеллы клетки, даже ядра. Поры играют существенную роль в переносе питательных веществ по гифам гриба в зону роста.
Настоящие ткани у аскомицетов формируются редко. Исключение составляют высокоспециализированные паразиты насекомых из порядка лабульбениевые – Laboulbeniales., вегетативное тело которых – рецептакул состоит из настоящей ткани. Основные полисахариды, входящие в состав клеточной стенки аскомицетов – хитин и гкюканы. Содержание хитина составляет 20-25%.
Размножение
В цикле развития многих аскомицетов большую роль играет бесполое размножение. Споры бесполого размножения – конидии – образуются на гаплоидном мицелии экзогенно (реже эндогенно) на конидиеносцах разного строения. Конидиеносцы образуются на мицелии одиночно, соединяются в пучки (коремии) или подушечки (спородохии), развиваются плотным слоем на поверхности сплетения гиф (ложа) или внутри шаровидных или грушевидных структур с отверстием на вершине пикниды.
Конидиальные спороношения развиваются в период вегетации грибов и служат для их массового расселения. У аскомицетов-паразитов они обычно образуются на живых растениях, а сумчатые спороношения (за немногими исключениями) – после отмирания растения или его частей в конце вегетации или после перезимовки.
У некоторых аскомицетов конидиальное спороношение неизвестно, у других оно преобладает в цикле развития. В отдельных группах этого класса сумчатая стадия образуется редко. Иногда её трудно обнаружить в природе и получить в искусственной культуре грибов, поэтому многие аскомицеты как в природе, так и в коллекциях культур чаще встречаются в конидиальной стадии.
 Рис.1. Половой процесс и развитие сумок у аскомицетов
Рис.1. Половой процесс и развитие сумок у аскомицетов
1- аскогон с трихогиной и антередий; 2 - развитие сумок по способу крючка; 3 - молодая сумка после мейоза; 4 - молодая сумка с восемью гаплоидными ядрами; 5 - зрелая сумка с аскоспорами
Конидиальные стадии многих аскомицетов имеют самостоятельные видовые наименования, нередко более распространенные, чем названия сумчатых стадий. Например, многие пенициллы и аспергиллы, образующие сумчатые стадии, более известны под названиями их конидиальных стадий. Так, широко используемый в генетических исследованиях аскомицет эмерицелла лежачая – Emericella nidulans - обычно называют аспергиллом лежачим - Aspergillus nidulans. В международном кодексе ботанической номенклатуры, который регулирует употребление названий растений, содержится специальный пункт, разрешающий использовать наряду с основным видовым названием аскомицета по его сумчатой стадии название его конидиальной стадии в тех случаях, когда речь идёт именно об этой стадии гриба.
Типичный для аскомицетов половой процесс - слияние двух специализированных клеток мицелия, не дифференцированных на гаметы. Такие клетки обычно называют гаметангиями, а тип полового процесса – гаметангиогамией. Однако «гаметангии» аскомицетов не гомологичны настоящим гаметангиям, в которых развиваются гаметы, а происходят, вероятно, от недифференцированных копулирующих ветвей мицелия соматогамных грибов.
У низших аскомицетов (подкласс гемиаскомицеты – Hemiascomycetidae) половой процесс сходен с зигогамией зигомицетов. Гаметангии разного пола морфологически сходны или малоразличимы и представляют выросты или веточки мицелия. После их слияния сразу наступает кариогамия, и сумка развивается непосредственно из зиготы. Однако в отличие от большинства зигомицетов в многоядерных гаметангиях аскомицетов сливаются только два ядра. Зигота не переходит в состояние покоя, а сразу развивается в сумку. В цикле развития низших аскомицетов имеются, таким образом, только гаплоидная и диплоидная фазы.
У высших аскомицетов (подклассы эуаскомицеты - Euascomycetidae и локулоаскомицеты - Loculofscomycetidae) наблюдается дифференциация и усложнение строения гаметангиев. Женский гаметангий состоит из двух частей – аскогона и нитевидно вытянутой трихогины, мужской гаметагий – антеридий – одноклеточный. При слиянии содержимое антеридия по трихогине переходит в аскогон. После плазмогамии гаплоидные ядра разного пола сразу не сливаются, а объединяются попарно, образую дикарион. Из аскогона вырастают аскогенные гифы, в которых ядра дикариона синхронно делятся. Эти гифы ветвятся и разделяются септами на двуядерные клетки. На аскогонных гифах развиваются сумки. Конечная клетка такой гифы разгибается крючком, ядра дикариона располагаются в месте перегиба и одновременно делятся. Пара ядер разного пола остаётся в месте перегиба крючка, одно ядро переходит в его кончик, еще одно – в основание. Затем образуются две перегородки, отделяющие одноядерные конечную и базальную клетки крючка. В результате слияния этих клеток восстанавливается дикарион и крючок сформироваться повторно. Средняя двуядерная клетка крючка развивается в сумку. Сумка увеличивается в размерах, ядра дикариона сливаются, диплоидное ядро делится редукционно и митотически и вокруг восьми гаплоидных ядер формируются аскоспора.
Типичный способ развития сумок был впервые описан П. Классеном у дискомицета пиронема омфалодес – Pyronema omphalodes в 1907 и 1912 гг. У некоторых аскомицетов (например, в порядке эвроциевые – Eurotiales) сумки образуются на аскогенных гифах иными способами: цепочками по ходу аскогенных гиф, из их конечных клеток или боковых выростов.
По способу формирования аскоспоры аскомицетов отличаются от спорангиоспор зародышевого спорангия зигомицетов. Если при образовании спорангиоспор происходит раскалывание цитоплазмы спорангия, то аскоспоры развиваются по способу так называемого «свободного образования клеток» - часть цитоплазмы сумки обособляются вокруг ядер и одеваются оболочкой. При этом аскоспоры и гемиаскомицетов и высших аскомицетов формируются по-разному: у первых наблюдается индивидуальное отделение каждой аскоспоры, у последних аскоспоры обособляются в результате инвагинации первоначально общей мембраны. В сумке аскоспоры окружены неиспользованной на их формирование цитоплазмы – эпиплазмой. К моменту созревания аскоспор в цитоплазме происходит превращение гликогена в сахар, тургорное давление в сумке резко возрастает, и аскоспоры с силой выбрасываются на расстояние от доли мм до 10см и более.
В результате образования аскогенных гиф увеличивается число сумок, а следовательно, и аскоспор, развивающихся из одного аскогона. Формирование сумок по способу крючка с восстановлением дикариона в его базальной клетке и повторным образованием крючков обеспечивает расположение сумок пучком или слоем, что облегчает активное освобождение аскоспор.
В цикле развития высших аскомицетов чередуется, таким образом, три фазы: длительная – гаплоидная, в течение которой происходит бесполое размножение, непродолжительная – дикариотическая (аскогенные гифы) и очень короткая – диплоидная (молодая сумка с диплоидным ядром).
Для многих аскомицетов характерна морфологическая редукция полового процесса. У некоторых представителей этого класса антеридии отсутствуют или не функционируют. В этом случае их функции могут выполнять конидии, вегетативные гифы или мелкие специализированные клетки, называемые спермациями. Спермации часто образуются на другом мицелии, на значительном расстоянии от аскогона и переносятся токами воздуха, каплями дождя, насекомыми. У некоторых аскомицетов трихогина хемотропична и подрастает к спермациям или конидиям совместимого типа.
В том случае, когда отсутствуют оба гемотангия, дикарионтизация происходит в результате слияния клеток вегетативных гиф одного или двух совместимых мицелиев – соматогамии. Иногда дикарионы формируются без слияния клеток – в результате попарной ассоциации ядер в аскогинии или в вегетоклетках гифы.
Строение оболочки
По строению оболочки и функциям сумки аскомицетов делят на две большие группы: прототуникатные и эутуникатные. Прототуникатные сумки имеют тонкую не дифференцированную оболочку, которая разрушается или растворяется, пассивно освобождая аскоспоры. Такая сумка служит только местом формирования спор, но активно не участвует в их распространении. Эутуникатные сумки характеризуются более плотными оболочками, часто со специальными приспособлениями для вскрывания сумки. Они активно участвуют в распространении аскоспор. Строение оболочки эутуникатных сумок может быть двух типов: уникатные и битуникатные.
1. Оболочка у унитуникатных сумок относительно тонкая и выглядит однослойной, на вершине сумок обычно имеет апикальный аппарат различного строения, служащий для их вскрывания.
2. Битуникатные сумки имеют ясно двуслойную оболчку, состоящую из жесткого наружного и эластичного внутреннего слоёв. При созревании аскоспор наружный слой оболочки разрушается, начиная с вершины, внутренний слой под действием повешенного тургорного давления растягивается, и аскоспоры активно выбрасываются.
У гемиаскомицетов и аскомицетов с замкнутыми плодовыми телами форма сумок округлая или овальная, у аскомицетов с активным выбрасыванием аскоспор – булавовидная или цилиндрическая.
В сумках аскомицетов обычно формируется 8 аскоспор. Однако наблюдаются многочисленные отклонения от типа. Сокращение числа аскоспор происходит в результате уменьшения числа деления ядер (например, образование четырёх аскоспор у эндомицеса Магнуса) или в результате дегенерации части гаплоидных ядер (например, развитие двух аскоспор у сморчковой шапочки). Число аскоспор увеличивается или при возрастании числа деления ядер в сумке, или в результате почкования аскоспор в сумке.
Форма аскоспор очень разнообразна от шаровидных или эллипсоидальных до нитевидных. Могут быть одноклеточными или могут иметь поперечные перегородки, реже – поперечные и продольные перегородки (муральные аскоспоры). У некоторых аскомицетов аскоспоры имеют разнообразные придатки, играющие роль в их распространении, например слизистые у копрофильных грибов. У низших аскомицетов сумки образуются непосредственно на мицелии, а у высших – в специальных вместилищах – плодовых телах и аскостромах.
Плодовые тела аскомицетов
Различают следующие типы плодовых тел: клейстотеций – полностью замкнутое плодовое тело, перитеций – полузамкнутое, обычно кувшиновидное плодовое тело с отверстием на вершине, апотеций – открытое, обычно чашевидное плодовое тело, на верхней стороне которого расположен слой сумок и парафиз. Клейстотеции, перитеции, апотеции – настоящие плодовые тела, развивающиеся по аскогеминиальному типу. Перидий (оболчка) такого плодового тела образуется после плазмогамии. Гаплоидные вегетативные гифы оплетают аскогенные гифы и сумки, формирую плотную покровную ткань.
 Рис.2. Плодовые тела эуаскомицетов
Рис.2. Плодовые тела эуаскомицетов
1,2 - клейстотеции; 3 - перитеций; 4,5 - апотеций; 6 - сумка; 7 - парафизы; 8 - перефизы
Настоящие плодовые тела могут развиваться как непосредственно на мицелии, так и на стромах – плотных сплетениях гиф различной формы, размера, консистенции. Аскострома развивается иначе. Сначала складывается строма из переплетающихся гиф. В ней образуются аскогоны и происходит половой процесс. Аскогонные гифы и появляющиеся на них сумки раздвигают или разрушают плектенхиму стромы, освобождая в ней полость (локулу). Каждая локула содержит одну или несколько сумок. Строма под локулой разрушается и образуется отверстие, через которое освобождаются аскоспоры.
Классификация аскомицетов
На основе отсутствия или наличия плодовых тел и их типа класс Ascomycetes делят на три подкласса:
1. Подкласс гемиаскомицеты, или голосумчатые – Hemiascomycetidae, - плодовые тела отсутствуют, сумки образуются непосредственно на мицелии, прототуникатные
2. Подкласс эуаскомицеты – Euascomucetidae – сумки образуются в клейстотециях, перитециях или апотециях, прототуникатные или уникатные.
3. Подкласс лакулоаскомицеты – Loculoascomucetidae – сумки образуются в аскостромах, битуникатные.
У подкласса эуаскомицетов (Euascomycetidae) сумки образуются в настоящих плодовых телах, развивающихся по аскогимениальному типу. Лишь у немногих примитивных представителей этого подкласса сумки формируются группами или пучками на мицелии и не окружены перидием или перидий плодового тела представляет рыхлое сплетение гиф, сквозь которое просвечивают сумки.
Система эуаскомицетов основана на строении плодовых тел, типе развития центра перитеция (у пиреномицетов), строении сумок, наличии и характере строме. В современной системе эуаскомицеты для удобства часто подразделяют на группы порядков соответственно типу плодовых тел и строению сумок, обычно не присваивая этим группам ранга таксонов.
К подклассу эуаскомицеты относят сейчас около двадцати порядков. Одна их групп порядков эуаскомицетов – группа порядков плектомицеты. К этой группе относят эуаскомицеты с клейстотециями, реже с перитециями, в которых беспорядочно располагаются прототуникатные сумки. Аскоспоры у этой группы всегда освобождаются пассивно. Одним из основных порядков этой группы является порядок эуроциевые – Eurotiales.
biofile.ru


















 |  |  |  |  |  |  |  |  |  |  |  |  |  |  |
 Пример видео 3 Пример видео 3 |  Пример видео 2 Пример видео 2 |  Пример видео 6 Пример видео 6 |  Пример видео 1 Пример видео 1 |  Пример видео 5 Пример видео 5 |  Пример видео 4 Пример видео 4 |
Администрация муниципального образования «Городское поселение – г.Осташков»