
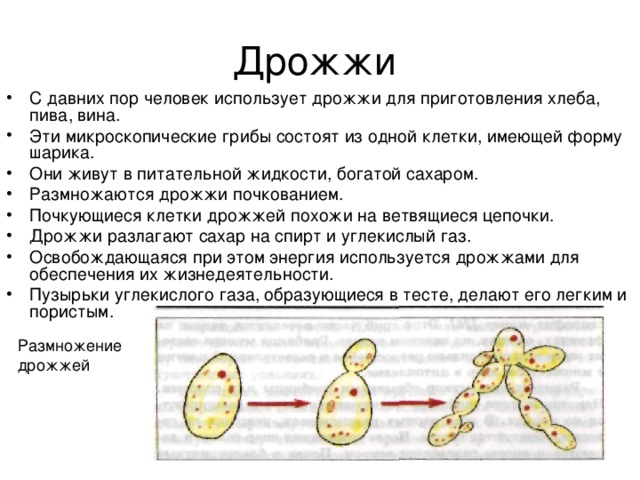
Содержание
Олимпиадные задания для 9 класса по биологии
Задания муниципального этапа олимпиады по биологии
9 класс – 2016 г
(макс. кол-во баллов 85,5.5)
Часть I. Вам предлагаются тестовые задания, требующие выбора только одного ответа из четырех возможных. Максимальное количество баллов, которое можно набрать – 40 (по 1 баллу за каждое тестовое задание). Индекс ответа, который вы считаете наиболее полным и правильным, укажите в матрице ответов.
1. На агар-агаре можно вырастить культуру возбудителей:
а) дизентерии
б) гриппа
в) малярии
г) диабета
2.Половое размножение хламидомонады происходит:
а) при достижении клетки достаточных размеров
б) при установлении определенного соотношения
между объемом ядра и цитоплазмы
в) при неблагоприятных условиях среды
г) при благоприятных условиях среды
3. Ложный опенок отличается от съедобного тем, что:
Ложный опенок отличается от съедобного тем, что:
а) ложный опенок намного крупнее
б) у ложного опенка нет пленки на пеньке
в) ложный опенок – трубчатый гриб
г) у ложного опенка светло-желтые пластинки
4. Малярийный плазмодий относится к типу:
а) саркодовых
б) жгутиковых
в) споровиков
г) инфузории
5. К одноклеточным организмам относятся грибы:
а) шампиньон и сыроежка;
б) микроспоридии и дрожжи;
в) пеницилл и мукор;
г) мухомор и дрожжи.
6. Корневые волоски – это
а) многоклеточные образования ризодермы
б) выросты клеток ризодермы
в) формирующиеся корневые клубеньки
г) очень маленькие по размеру боковые корни
7. У василька синего цветки:
а) язычковые и трубчатые
б) трубчатые и воронковидные
в) язычковые и воронковидные
г) ложноязычковые и трубчатые
8. Центральный цилиндр корня отделяется от первичной коры клетками:
а) мезодермы
б) перицикла
в) эндодермы
г) эктодермы
9. Какой из перечисленных газов является гормоном растений?
Какой из перечисленных газов является гормоном растений?
а) ацетилен
б) этилен
в) пропан
г) углекислый газ
10. Пыльцевход находится
а) на рыльце пестика
б) в семязачатке
в) в зародышевом мешке
г) в тычинках
11.Запасные белки у растений накапливаются в:
а) бесцветных пластидах
б) хлоропластах в) клеточном соке
г) вакуолях
12. Основное значение фотолиза воды в процессе фотосинтеза — это:
а) восполнение недостающего электрона в пигменте реакционного центра
б) выделение кислорода растениями в атмосферу Земли
в) образование метаболической воды
г) образование как можно большего количества протонов внутри тилакоидов
13. В каком из примеров правильно отражена последовательная смена растений при первичной сукцессии?
а) мхи – лишайники – травянистые растения
б) лишайники – травянистые растения – мхи
в) лишайники – мхи – травянистые растения
г) травянистые растения – мхи – лишайники
14. Концепцию понятия «биосфера» впервые предложил :
Концепцию понятия «биосфера» впервые предложил :
а) Константин Эдуардович Циолковский
б) Владимир Николаевич Сукачев
в) Жан Батист Ламарк
г) Владимир Иванович Вернадский
15. У высших растений в состав клеточной стенки входит(-ят)
а) только целлюлоза
б) только целлюлоза и пектины
в) целлюлоза и хитин
г) целлюлоза, гемицеллюлозы, пектины, белки
16. У кишечнополостных между эктодермой и энтодермой находится:
а) полость тела
б) мезодерма
в) мезоглея
г) кишечная полость
17. Личинки каких насекомых лишены членистых конечностей?
а) чешуекрылых
б) жесткокрылых
в) полужесткокрылых
г) двукрылых
18. Что такое регенерация?
а) восстановление утраченных частей тела
б) бесполый способ размножения животных
в) половой способ размножения животных
г) защита от нападения
19. Пара животных с одинаковым числом усиков:
Пара животных с одинаковым числом усиков:
а) водомерка и тутовый шелкопряд
б) блоха и речной рак
в) паук-крестовик и речной рак
г) тигровая креветка и собачий клещ
20. Прудовики дышат
а) жабрами
б) трахеями;
в) легкими;
г) поверхностью тела
21. Рептилии, как и птицы,
а) откладывают яйца, покрытые твёрдой известковой оболочкой
б) имеют сухую кожу без желёз
в) способны поддерживать постоянную температуру тела
г) имеют лёгкие, неспособные к растяжению
22. Какие плавники играют ведущую роль при движении рыбы вперед в открытой воде?
а) грудные
б) брюшные
в) спинной
г) хвостовой
23. Какая группа является наиболее древней среди современных рептилий:
а) крокодилы
б) черепахи
в) гаттерии
г) змеи
24. У птиц киль служит для:
а) рассекания воздуха
б) устойчивости в полете
в) прикрепления мышц
г) защиты внутренних органов
25. Человеческий плод человека соединяется с материнским организмом через:
Человеческий плод человека соединяется с материнским организмом через:
а) плаценту
б) стенку матки
в) пуповину
г) желтое тело
26. К отряду Перепончатокрылые относятся:
а) пчела, оса, овод
б) шершень, наездник, муравей
в) шмель, слепень, богомол
г) пилильщик, рогохвост, стрекоза
27. Из перечисленных объектов, к первичноводным животным относится:
а) крокодил
б) дельфин
в) ихтиозавр
г) тритон
28. Отличительным признаком между твердой и мягкой пшеницей является то, что :
а) у твердой пшеницы выше содержание белка
б) твердая пшеница более устойчива к холоду
в) твердую пшеницу можно сеять позже мягкой
г) из твердой пшеницы получают более дешевую муку
29. Соединение ребер с позвонками у человека:
а) подвижное
б) полуподвижное
в) неподвижное
г) вообще не соединяются
30. Что произойдёт с озимыми злаками, если высадить их весной одно- временно с яровыми?
а) Они взойдут и дадут урожай быстрее яровых
б) Они образуют вегетативные органы, но не перейдут к цветению
в) Они взойдут и засохнут из-за недостатка влаги и высоких температур
г) Они ничем не будут отличаться от яровых
31. Синтез белка не происходит в следующих органоидах клетки:
Синтез белка не происходит в следующих органоидах клетки:
а) рибосомах
б) лизосомах
в) митохондриях
г) ЭПР
32. ДНК бактерий состоит из:
а) одной цепи ДНК;
б) двух цепей ДНК;
в) белка и РНК;
г) четырех молекул ДНК
33.Рост организма человека регулируется следующими гормонами:
а) гормоном роста, тиреоидными гормонами, инсулином;
б) гормоном роста, вазопрессином, тиреоидными гормонами;
в) гормоном роста, антидиуретическим гормоном, пролактином;
г) гормоном роста, адреналином.
34. В обеспечении состояния бодрствования важнейшую роль играет
а) чёрная субстанция среднего мозга
б) ретикулярная формация ствола мозга
в) мозолистое тело
г) гиппокамп
35. Генетический материал вируса СПИДа представлен:
а) одноцепочечной ДНК;
б) двуцепочеченой ДНК;
в) одноцепочечной РНК;
г) двухцепочечной РНК.
36. Центриоли удваиваются в:
Центриоли удваиваются в:
а) G1-фазе клеточного цикла;
б) S-фазе клеточного цикла;
в) G2-фазе клеточного цикла;
г) процессе митотического деления.
37. В норме оплодотворённая яйцеклетка у плацентарных млекопитающих начинает делиться в
а) полости тела
б) яйцеводе
в) матке
г) яичнике
38. Антитела вырабатывают
а) эритроциты
б) Т-лимфоциты
в) В-лимфоциты
г) макрофаги
39. Паратгормон паращитовидной железы активизирует поглощение кальция из кишечника в кровь при условии достаточного поступления в организм человека витамина:
а) D;
б) С;
в) Е;
г) В1.
40. Аминокислота соединяется со своей тРНК посредством :
а) фермента аминоацил-тРНК-синтетазы без затраты АТФ;
б) фермента РНК-полимеразы без затраты АТФ;
в) фермента РНК-полимеразы с затратой АТФ;
г) фермента аминоацил-тРНК-синтетазы с затратой АТФ.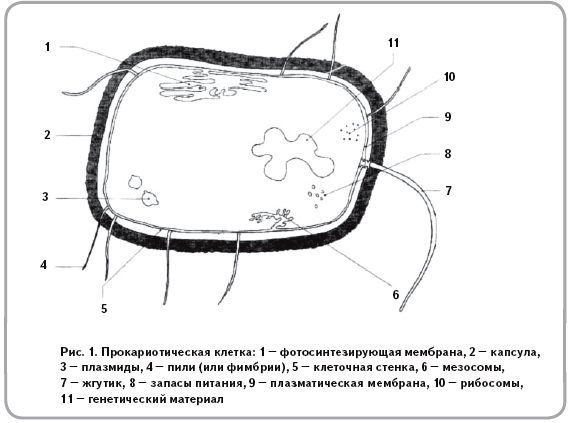
Часть 2. Вам предлагаются тестовые задания с одним вариантом ответа из четырёх возможных, но требующие предварительного множественного выбора. Максимальное количество баллов, которое можно – 20 (по 1 баллу за каждое тестовое задание). Индекс ответа, который вы считаете наиболее полным и правильным, укажите в матрице ответов.
1. Не имеют жесткой клеточной стенки клетки:
1. дрожжей;
2. костной ткани;
3. листа березы;
4. амёбы;
5. туберкулезной палочки.
а) 1,2,4;
б) 2,4;
в) 1,3,5;
г) 2,4,5;
д) 1,3.
2.Бактерии вызывают заболевания:
1) малярия
2) сыпной тиф
3) столбняк
4) дизентерия
5) гепатит
а) 2, 3, 4;
б) 1, 3, 5;
в) 1, 2, 4;
г) 2, 4, 5;
д) 2, 3, 4, 5.
3.Главный корень покрытосеменного растения может:
1) поглощать воду с растворенными веществами;
2) закреплять растения в почве;
3) расти за счет вставочной меристемы;
4) синтезировать аминокислоты, гормоны, алкалоиды;
5) образовывать чешуевидные листья на старых участках корней.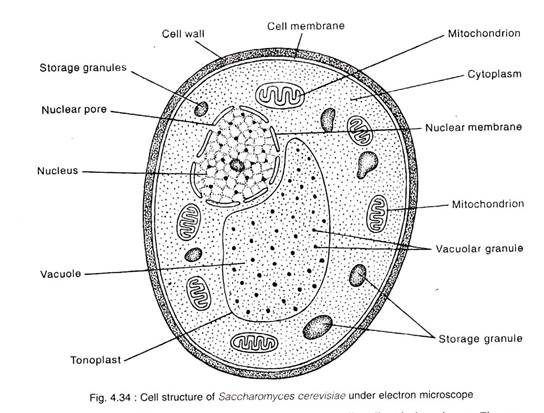
а) 1, 2, 3;
б) 1, 2, 4;
в) 2, 4, 5;
г) 2, 3, 4
д) 1, 4,
4.Из названных веществ нейромедиаторами ЦНС являются:
1) адреналин;
2) норадреналин;
3) ацетилхолин;
4) вещество Р;
5) глутамат.
а) 1, 2, 3, 4;
б) 2, 3, 4, 5;
в) только 2, 4, 5;
г) только 1, 4, 5.
д)только 4,5.
5. Из перечисленных животных трахейную систему имеет:
1) паук крестовик;
2) сольпуга;
3) скорпион;
4) мокрица;
5) перипатус.
а) только 1, 2, 3;
б) только 2, 4;
в) только 1, 3;
г) 1, 2, 4, 5
д) только 2,3.
6. Из приведенных черт ехидны, характерными только для класса млекопитающих являются:
1) часть ее тела покрыта волосами;
2) полное разделение малого и большого круга кровообращения и четырехкамерное сердце;
3) грудная и брюшная полости тела разделены мышечной диафрагмой.
4) температура тела не зависит от температуры окружающей среды;
5) эритроциты безъядерные.
а) 1, 3, 5;
б) 2, 3; 4;
в) 1, 3, 4;
г) 2, 3, 5;
д) 1,2,5.
7. К моносахаридам относятся углеводы:
1) рибоза;
2) глюкоза;
3) целлюлоза;
4) фруктоза;
5) крахмал
а) 1, 2,3;
б) 1,2,4;
в) 1, 3, 5;
г) 2, 3, 5; д) 1,4,5.
8. Орган/органы, в котором капиллярную сеть образуют не артерии, а вены:
1) конечный мозг;
2) сердце;
3) желудок;
4) печень;
5) пищевод.
а) 1, 2;
б) 3, 4;
в) только 1;
г) только 4;
д)только 5 .
9. Из средиземноморского центра происхождения культурных растений по Н.И.Вавилову) были введены в культуру:
1) капуста;
2) картофель;
3) ячмень;
4) петрушка;
5) свёкла а) 1, 2, 4; б) 2, 3, 4; в) 1, 4, 5 г) 2, 3, 5; д) 3,4,5.
10. В состав лицевого черепа входят следующие кости:
1) решетчатая;
2) носовая;
3) верхнечелюстная;
4) сошник;
5) клиновидная.
а) 1, 2, 3,4;
б) 2;3,4;
в) 1, 4, 5;
г) 2, 3, 4;
д) 2,3,5.
Часть 3. Вам предлагаются тестовые задания в виде суждений, с каждым из которых следует либо согласиться, либо отклонить. В матрице ответов укажите вариант ответа «да» или «нет». Максимальное количество баллов, которое можно набрать – 15 (по 1 баллу за каждое задание).
1.Удаление плодового тела трутовика со ствола дерева избавляет растение от паразита.
2. Земноводные получают кислород через лёгкие и кишечник.
3. Яйцекладущие млекопитающие встречаются в Австралии и Южной Америке.
4. Орангутанги являются ближайшими родственниками человека .
5. Самым протяженным отделом пищеварительной системы является толстый кишечник .
6. Двоякодышащие рыбы – вымершая группа рыб, от которой произошли первые земноводные.
7. Плацента может выполнять секреторную функцию как железа внутренней секреции .
8. Женские особи медицинских пиявок (Hirudo medicalis)крупнее мужских .
9. Дифференцировка всех лимфоцитов происходит в тимусе.
10. Наибольшая скорость движения крови наблюдается в полых венах.
11. Эпителиальные ткани делят на две группы: покровные и железистые .
12. Эндосперм покрытосеменных растений является видоизмененным женским гаметофитом.
13. Структурная единица почки человека – нефрон – образован из мальпигиевого клубочка.
14. Сукцессия возникает в результате нарушения равновесия в экосистеме.
15. Желтое тело сохраняется, если оплодотворенная яйцеклетка имплантируется в эндометрий.
Часть 4. Вам предлагаются тестовые задания, требующие установления соответствия. Максимальное количество баллов, которое можно набрать – 10,5. Заполните матрицы ответов в соответствии с требованиями заданий.
1. [мах. 2,5 балла] Соотнесите систематические группы растений (А–Б) с их
признаками (1–5).
1. Гаметофит раздельнополый. |
А. Покрытосеменные |
Признаки | 1 | 2 | 3 | 4 | 5 |
Систематическая группа |
|
|
|
|
|
2. [мах. 4 балла] Установите соответствие между конечностями речного рака (1 – 8) и функциями, которые они выполняют (А – З) Функции конечностей:
А) обеспечивают плавание головой вперёд
Б) перетирают пищу
В) измельчают пищу и транспортируют её ко рту, участвуют в газообмене
Г) направляют пищу ко рту, гонят воду через жаберную полость
Д) обеспечивают плавание задом наперёд
Е) обеспечивают передачу спермы при спаривании
Ж) обеспечивают передвижение по дну, участвуют в захвате пищи и газообмене
З) сенсорные придатки
Конечности | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
Функции |
|
|
|
|
|
|
|
|
3. [мах. 4 балла] Соотнесите характеристику (1 – 4) с типом межклеточной сигнализации (А–В), к которому она относится.
Характеристика: | Тип сигнализации |
1) Действие сигнального вещества направлено на ту клетку, которая его выделяет. |
А. Аутокринный
|
Характеристика: | 1 | 2 | 3 | 4 |
Тип сигнализации |
|
|
|
|
Ответы на задания муниципального этапа олимпиады
по биологии 9 класс
Часть 1. [40 баллов]
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
1-10 | а | в | б | в | б | б | б | б | б | б |
11-20 | в | а | в | в | г | в | г | а | а | в |
21-30 | а | г | в | в | в | б | г | а | а | б |
31-40 | б | а | а | б | в | б | б | в | а | г |
Часть 2 [20 баллов]
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
1-10 | б | а | б | б | г | а | б | г | в | б |
Часть 3 [15 баллов]
№ | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
Да |
| х | х |
|
| х |
|
|
| |
Нет | х | х |
| х | х |
| х | х | х | |
| ||||||||||
| 11 | 12 | 13 | 14 | 15 |
|
|
|
|
|
Да | х |
| х | х |
|
|
|
|
| |
Нет |
| х | х |
|
|
|
|
|
|
|
Часть 4 [10,5 баллов]
1. [мах. 2,5 балла]
Признаки | 1 | 2 | 3 | 4 | 5 |
Систематическая группа | а | б | б | б | а |
2. [мах. 4 баллов]
Конечности | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
Функции | в | г | е | а | ж | з | б | д |
3. [мах. 4 баллов]
Характеристика: | 1 | 2 | 3 | 4 |
Тип сигнализации | а | в | б | в |
Как археи «толстеют» из-за вирусов
27 апреля 2021
Вирусы архей — пожалуй, самые загадочные и малоизученные вирусы, не имеющие эволюционного родства ни с вирусами бактерий, ни с вирусами эукариот. Почти в каждой посвященной им работе обнаруживаются их уникальные свойства, не описанные ни у каких других вирусов. В недавней статье, опубликованной в PNAS, сообщается, что вирус STSV2, поражающий архей рода Sulfolobus, до неузнаваемости изменяет морфологию инфицированных клеток: они увеличиваются в размере почти в 20 раз и от бинарного деления переходят к почкованию, похожему на почкование дрожжей. Почему так происходит? Давайте разбираться.
Многие вирусы научились перестраивать под себя нормальный клеточный цикл клетки-хозяина. Например, некоторые вирусы эукариот стимулируют переход клеток из фазы G1 клеточного цикла в S-фазу, чтобы удваивать свои геномы вместе с геномом клетки-хозяина. Клеточный цикл архей чем-то напоминает таковой у эукариот, хоть и с некоторыми отличиями. Лучше всего он изучен у архей рода Sulfolobus — экстремофилов, обитающих при температуре около 80 °C и pH 3. В экспоненциально растущих клетках Sulfolobus выделяют пререпликативный период (G1-фаза), стадию репликации хромосомы (S-фаза), второй период клеточного роста (G2-фаза), быструю сегрегацию геномов и собственно разделение дочерних клеток, обозначаемые как M- и D-фазы соответственно. Ключевой фактор деления бактериальных клеток близок к эукариотическому комплексу ESCRT (от англ. endosomal sorting complex required for transport), который играет важную роль в перестройках мембран во время формирования везикул [1].
Именно у гипертермофильных архей описано множество разнообразных вирусов .
Подробнее о вирусах архей вы можете прочитать в нашей статье «Загадочные вирусы архей» [2].
В отличие от бактериофагов, которые зачастую быстро убивают клетку, вирусы архей, как и вирусы эукариот, чаще превращают инфицированную клетку в настоящую фабрику по производству вирусных частиц. Иногда такие клетки называют вироклетками. Чтобы сделать из клетки комбинат по производству вирусных частиц, вирусы хитрым образом перестраивают клеточный цикл и другие аспекты функционирования клетки под себя, изменяя ее до неузнаваемости [1].
Авторы недавнего исследования, опубликованного в PNAS, изучали взаимодействие клеток архей и вирусов на примере нелитического, то есть не вызывающего быструю гибель клетки, вируса STSV2 семейства Bicaudaviridae и его хозяина — гипертермофильной археи Sulfolobus islandicus REY15A. Ученые наблюдали за динамикой зараженной и незараженной клеточных популяций в течение 10 дней, измеряя оптическую плотность культуры. Поначалу плотность зараженной культуры, как и следовало ожидать, снижалась вследствие замедления роста и гибели клеток, а титр вирусных частиц в ней возрастал. Однако к 7 дню после заражения ситуация начала меняться: плотность инфицированной популяции начала быстро расти, а титр вирусных частиц, наоборот, стал снижаться, что свидетельствует о появлении в ней клеток, устойчивых к вирусу [1].
Чтобы детально изучить особенности клеток на разных стадиях инфекции, авторы брали образцы зараженной культуры спустя разное время после инфицирования и визуализировали клетки с помощью световой микроскопии (подробнее о методах микроскопии читайте в статье «12 методов в картинках: микроскопия» [3]). Они обнаружили удивительный факт: клетки, инфицированные STSV2, значительно увеличивались в размерах. Если на первые сутки после инфицирования они превосходили здоровые клетки по размеру в два раза, то постепенно эта разница увеличивалась аж до 20 раз к шестому дню после инфицирования! Изучение зараженных клеток с помощью сканирующей электронной микроскопии позволило установить, что их поверхность густо усеяна вирионами STSV2. Интересно, что, начиная с седьмого дня после инфицирования, доля клеток нормального размера постепенно начинала возрастать, и после восьмого дня инфекции уже 96% клеток имели нормальные размеры (рис. 1) [1].
Рисунок 1. Динамика изменений размеров клеток в инфицированной и неинфицированной популяциях (dpi — день после заражения). а — Световая микроскопия. б — Сканирующая электронная микроскопия. На рисунке а масштабная линейка 2 мкм, на рисунке б — 1 мкм.
Ученые также исследовали, как меняется количество ДНК в популяции зараженных клеток архей. В обычной популяции Sulfolobus islandicus большинство клеток находятся в фазе G2 и содержат две копии хромосомы, а меньшая доля клеток, находящаяся в фазе S, имеет одну хромосому. По мере приближения к стационарной фазе роста в популяции начинают преобладать клетки в G1-фазе с одной хромосомой. Авторы работы исследовали количество ДНК в клетках с помощью проточной цитофлуориметрии и выяснили, что спустя сутки после заражения около 80% клеток содержали более четырех копий хромосом. По мере развития инфекции количество ДНК в клетках увеличивалось, и в некоторых зараженных клетках достигало целых 300 копий хромосомы. В то же время, начиная с шестых суток после заражения, в популяции стали выявляться клетки, содержащие одну или две копии хромосомы, что согласуется с наблюдениями, полученными с помощью микроскопии: в какой-то момент на лидирующее положение в популяции вместо клеток-гигантов начинали выходить клетки нормального размера [1].
О проточной цитофлуориметрии можно прочитать в статье «12 методов в картинках: проточная цитофлуориметрия» [4].
А что же происходит в ходе инфекции с количеством вирусной ДНК? Для ответа на этот вопрос ученые брали пробы инфицированных клеток на разных стадиях инфекции, выделяли из них ДНК и с помощью ПЦР в реальном времени определяли принадлежность ДНК — является ли она вирусной или клеточной (подробнее про ПЦР читайте в статье «12 методов в картинках: полимеразная цепная реакция» [5]). Здесь исследователей снова ждал сюрприз. В ходе инфекции количество вирусных геномов в зараженных клетках стремительно росло и к шестому дню достигало более двух тысяч копий на одну гигантскую клетку, однако после этого число вирусных геномов резко шло на спад, что также согласуется с гипотезой о том, что после шестого дня инфекции в популяции начинают доминировать резистентные клетки нормального размера. Стоит отметить, что вирусные геномы в инфицированных клетках равномерно распространены по цитоплазме и не образуют выраженных скоплений [1].
Но почему же зараженные клетки превращаются в гигантских полиплоидных монстров? Самая очевидная гипотеза — вирус каким-то образом препятствует финальной стадии деления, то есть разделению дочерних клеток. И действительно, анализ экспрессии генов компонентов ESCRT-подобного комплекса показал, что в инфицированных клетках уровень его синтеза сильно снижен по сравнению с нормальными и достигает самых низких значений на второй день инфекции. В период с 3 по 6 день инфекции уровень экспрессии генов ESCRT в зараженных клетках остается постоянным, а после шестого дня инфекции, когда в популяции начинают доминировать нормальные клетки, он поднимается до значений, характерных для неинфицированных клеток. Одновременно с повышением уровня производства комплекса ESCRT растет и оптическая плотность культуры, что указывает на интенсивное деление клеток [1].
На протяжении всего эксперимента, помимо гигантских клеток, примерно 20% популяции составляли клетки нормального размера. Откуда они берутся? Детальное исследование гигантских клеток показало наличие на их поверхности бугорков, которые по размерам соответствовали нормальным клеткам архей. Авторы работы предположили, что нормальные клетки отпочковываются от гигантских в ходе процесса, похожего на почкование дрожжей. Действительно, иммунофлуоресцентный анализ показал, что в местах отхождения бугорков от клеточной поверхности образуется кольцо или, точнее, спиральная сеть из комплекса ESCRT, который в нормальных клетках локализуется в их центре и обеспечивает равное бинарное деление (рис. 2) [1].
Рисунок 2. «Почкование» инфицированных архей. Зеленым цветом окрашены мембраны; синим — ДНК; красным — компонент ESCRT. Изображения в двух правых колонках получены наложением изображений, снятых в разных каналах. Видно, что ESCRT словно обвивает спиралью зарождающуюся почку.
Как мы отмечали ранее, в какой-то момент гигантские клетки почти полностью исчезают из популяции, и большинство вновь начинают составлять клетки нормального размера. Ученые показали, что эти клетки обладают устойчивостью к STSV2 благодаря системе CRISPR. Этап адаптации, то есть вставка спейсеров, соответствующих STSV2, начинается примерно на седьмой день после заражения, причем адаптация происходит не только в клетках нормального размера, но и в гигантских клетках [1].
STSV2 — не единственный вирус архей, который вызывает резкое увеличение зараженных клеток в размерах. Авторы показали, что схожие изменения вызывает вирус SMV1, который, как и STSV2, относится к семейству Bicaudaviridae, при этом вирус SSV2, принадлежащий к другому семейству, на размер инфицированных клеток не влияет. Авторы отмечают, что индукция гигантизма инфицированных архей может являться консервативной стратегией вирусов семейства Bicaudaviridae.
Примечательно также, что резкое увеличение размеров клеток архей возможно и по иным причинам. Например, архея Staphylothermus marinus, родственная Sulfolobus, при выращивании на среде, содержащей большое количество дрожжевого экстракта, может дорастать до диаметра 15 мкм при нормальном диаметре 0,5–1 мкм. Авторы исследования предполагают, что пластичность клеток архей в отношении размеров может быть обусловлена тем, что они, в отличие от большинства бактерий, не имеют жесткой пептидогликановой клеточной стенки [1].
- Junfeng Liu, Virginija Cvirkaite-Krupovic, Diana P. Baquero, Yunfeng Yang, Qi Zhang, et. al.. (2021). Virus-induced cell gigantism and asymmetric cell division in archaea. Proc Natl Acad Sci USA. 118, e2022578118;
- Загадочные вирусы архей;
- 12 методов в картинках: микроскопия;
- 12 методов в картинках: проточная цитофлуориметрия;
- 12 методов в картинках: полимеразная цепная реакция.

Архитектура клеточных стенок дрожжей: новая структура и новые задачи
1. Бартницки-Гарсия С., Перссон Дж., Чанзи Х. Исследование с помощью электронного микроскопа и электронной дифракции влияния калькофлуорового белого на биосинтез хитина in vitro . Арх Биохим Биофиз. 1994; 310:6–15. [PubMed] [Google Scholar]
2. Блэквелл Дж. Макромолекулярная организация целлюлозы и хитина. В: Браун Р. М. мл., редактор. Целлюлоза и другие природные полимерные системы. Нью-Йорк, Нью-Йорк: Plenum Press; 1982. стр. 403–428. [Google Scholar]
3. Bony M, Thines-Semproux D, Barre P, Blondin B. Локализация и закрепление на клеточной поверхности флоккуляционного белка Saccharomyces cerevisiae Flo1p. J Бактериол. 1997; 179:4929–4936. [Бесплатная статья PMC] [PubMed] [Google Scholar]
4. Cabib E, Bowers B, Roberts RL. Векторный синтез полисахарида изолированными плазматическими мембранами. Proc Natl Acad Sci USA. 1983; 80: 3318–3321. [Бесплатная статья PMC] [PubMed] [Google Scholar]
5. Кабиб Э., Дргон Т., Дргонова Дж., Форд Р.А., Коллар Р. Стенка дрожжевой клетки, динамическая структура, участвующая в росте и морфогенезе. Биохим Сок Транс. 1997; 25: 200–204. [PubMed] [Google Scholar]
6. Cappellaro C, Baldermann C, Rachel R, Tanner W. Специфическое для типа спаривания распознавание клеток Saccharomyces cerevisiae : прикрепление к клеточной стенке и активные участки a- и α-агглютинина . EMBO J. 1994; 13:4737–4744. [Бесплатная статья PMC] [PubMed] [Google Scholar]
7. Caro L H P, Tettelin H, Vossen J H, Ram A F J, van den Ende H, Klis F M. In silicio идентификация гликозил-фосфоатидилинозитол-заякоренных белков плазматической мембраны и стенок Saccharomyces cerevisiae . Дрожжи. 1997; 13:1447–1489. [PubMed] [Google Scholar]
8. Chen M-H, Shen Z-M, Bobin S, Kahn PC, Lipke PN. Структура Saccharomyces cerevisiae α-агглютинина: свидетельство наличия белка клеточной стенки дрожжей с множественными иммуноглобулиноподобными доменами. с атипичными дисульфидами. Дж. Биол. Хим. 1995; 270:26168–26177. [PubMed] [Академия Google]
9. Cid V J, Duran A, del Rey F, Snyder M P, Nombela C, Sanchez M. Молекулярные основы клеточной целостности и морфогенеза в Saccharomyces cerevisiae . Microbiol Rev. 1995; 59:345–386. [Бесплатная статья PMC] [PubMed] [Google Scholar]
10. Косгроув Д. Дж. Ползучие, стенки, размягчение плодов и проникающие пыльцевые трубки: растущая роль экспансинов. Proc Natl Acad Sci USA. 1997; 94: 5504–5505. [Бесплатная статья PMC] [PubMed] [Google Scholar]
11. De Nobel H, Lipke PN. Играют ли роль GPI в сборке клеточной стенки дрожжей? Тенденции клеточной биологии. 1994;4:42–45. [PubMed] [Google Scholar]
12. Де Нобель Дж. Г., Клис Ф. М., Прием Дж., Мунник Т., ван ден Энде Х. Растворимые в глюканазе маннопротеины ограничивают пористость клеточной стенки у Saccharomyces cerevisiae . Дрожжи. 1990; 6: 491–499. [PubMed] [Google Scholar]
13. Dijkgraaf GJP, Brown JL, Bussey H. Ген KNh2 Saccharomyces cerevisiae является функциональным гомологом KRE9 . Дрожжи. 1996; 12: 683–692. [PubMed] [Google Scholar]
14. Frevert J, Ballou C E. Saccharomyces cerevisiae структурный маннопротеин клеточной стенки. Биохимия. 1985; 24: 753–759. [PubMed] [Google Scholar]
15. Фрай С. С. Сшивание матричных полимеров в растущих клеточных стенках покрытосеменных растений. Annu Rev Plant Physiol. 1986; 37: 165–186. [Google Scholar]
16. Гатти Э., Пополо Л., Вай М., Рота Н., Альбергина Л. О-связанные олигосахариды в дрожжевом гликозилфосфатидилинозитол-заякоренном белке gp115 сгруппированы в богатой серином области, не существенной для его функции. Дж. Биол. Хим. 1994;269:19695–19700. [PubMed] [Google Scholar]
17. Gentzsch M, Tanner W. Протеин-O-гликозилирование в дрожжах: белок-специфические маннозилтрансферазы. Гликобиология. 1997; 7: 481–486. [PubMed] [Google Scholar]
18. Goldman RC, Sullivan PA, Zakula D, Copobianco JO. Кинетика взаимодействия β1,3-глюкана на донорных и акцепторных участках грибковой глюкозилтрансферазы, кодируемой геном BGL2 . Евр Дж Биохим. 1995; 227: 372–378. [PubMed] [Google Scholar]
19. Хэнсон Б.А., Лестер Р.Л. Влияние инозитолового голодания на in vitro синтез маннана и N-ацетилглюкозаминилпирофосфорилдолихола в Saccharomyces cerevisiae . J Бактериол. 1982; 151: 334–342. [Бесплатная статья PMC] [PubMed] [Google Scholar]
20. Hartland RP, Vermeulen CA, Klis FM, Sietsma JH, Wessels JG. Связь (1-3)-β-глюкана с хитином во время сборки клеточной стенки в Saccharomyces cerevisiae . Дрожжи. 1994; 10:1591–1599. [PubMed] [Google Scholar]
21. Horvath A, Sutterlin C, Manning-Krieg U, Movva NR, Riezman H. Синтез церамидов усиливает транспорт GPI-заякоренных белков в аппарат Гольджи у дрожжей. EMBO J. 1994;13:3687–3695. [Бесплатная статья PMC] [PubMed] [Google Scholar]
22. Джентофт Н. Почему белки O-гликозилированы? Тенденции биохимических наук. 1990; 15: 291–295. [PubMed] [Google Scholar]
23. Kaiser C, Gimeno R, Shaywitz D. Секреция белков, мембранный биогенез и эндоцитоз. В: Прингл Дж., Броуч Дж., Джонс Э., редакторы. Молекулярная и клеточная биология дрожжей Saccharomyces . 3. Клеточный цикл и клеточная биология. Колд-Спринг-Харбор, Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор; 1997. стр. 91–227. [Google Scholar]
24. Kapteyn J C, Montijn RC, Dijkgraaf G J, Van den Ende H, Klis F M. Ковалентная ассоциация β-1,3-глюкана с β-1,6-глюкозилированными маннопротеинами в клеточных стенках Кандида альбиканс . J Бактериол. 1995;177:3788–3792. [Бесплатная статья PMC] [PubMed] [Google Scholar]
25. Kapteyn J C, Montijn RC, Vink E, de la Cruz J, Llobell A, Douwes J E, Shimoi H, Lipke PN, Klis FM. Retention of Saccharomyces cerevisiae белков клеточной стенки через гетерополимер β-1,3-/β-1,6-глюкана, связанный фосфодиэфирной связью. Гликобиология. 1996;6:337–345. [PubMed] [Google Scholar]
26. Kapteyn J C, Ram A F, Groos E M, Kollar R, Montijn RC, Van Den Ende H, Llobell A, Cabib E, Klis FM. Измененная степень сшивки β1, 6-глюкозилированные маннопротеины в хитин у мутантов Saccharomyces cerevisiae со сниженным содержанием β1,3-глюкана клеточной стенки. J Бактериол. 1997; 179:6279–6284. [Бесплатная статья PMC] [PubMed] [Google Scholar]
27. Коллар Р., Петракова Э., Эшвелл Г., Роббинс П. В., Кабиб Э. Архитектура клеточной стенки дрожжей. Связь между хитином и β(1→3)-глюканом. Дж. Биол. Хим. 1995;270:1170–1178. [PubMed] [Google Scholar]
28. Коллар Р., Рейнхольд Б.Б., Петракова Е., Йе Х. Дж., Эшвелл Г., Дргонова Дж., Каптейн Дж. К., Клис Ф. М., Кабиб Е. Архитектура клеточной стенки дрожжей. β(1→6)-глюкан связывает маннопротеин, β(1→)3-глюкан и хитин. Дж. Биол. Хим. 1997; 272:17762–17775. [PubMed] [Google Scholar]
29. Копецка М., Крегер Д. Р. Сборка микрофибрилл in vivo и in vitro из 1-3β-D-глюкана, синтезированного протопластами Saccharomyces cerevisiae . Арка микробиол. 1986; 143: 387–395. [PubMed] [Google Scholar]
30. Копецка М., Фафф Х.Дж., Флит Г.Х. Демонстрация фибриллярного компонента в клеточной стенке дрожжей Saccharomyces cerevisiae и его химическая природа. Джей Селл Биол. 1974; 62: 66–76. [Бесплатная статья PMC] [PubMed] [Google Scholar]
31. Krainer E, Stark R E, Naider F, Alagramam K, Becker J M. Прямое наблюдение глюканов клеточной стенки в целых клетках Saccharomyces cerevisiae под магическим углом спиннинг 13 С-ЯМР. Биополимеры. 1994; 34: 1627–1635. [PubMed] [Google Scholar]
32. Лейдич С. Д., Костова З., Латек Р. Р., Костелло Л. С., Драпп Д. А., Грей В., Фасслер Дж. С., Орлеан П. Термочувствительные дрожжевые якорные мутанты GPI gpi2 и gpi3 нарушение синтеза N-ацетилглюкозаминилфосфатидилинозитола. Клонирование гена GPI2 . Дж. Биол. Хим. 1995; 270:13029–13035. [PubMed] [Google Scholar]
33. Лопес-Рибо Дж. Л., Аллуш Х. М., Мастен Б. Дж., Чаффин В. Л. Доказательства присутствия в клеточной стенке Candida albicans белка, относящегося к семейству hsp70. Заразить иммун. 1996;64:3333–3340. [Бесплатная статья PMC] [PubMed] [Google Scholar]
34. Lu C-F, Montijn RC, Brown JL, Klis F, Kurjan J, Bussey H, Lipke PN. Гликозилфосфатидилинозитол-зависимая перекрестная связь α-агглютинина и β1,6-глюкан в клеточной стенке S. cerevisiae . Джей Селл Биол. 1995; 128: 333–340. [Бесплатная статья PMC] [PubMed] [Google Scholar]
35. Lussier M, Sdicu AM, Winnett E, Vo DH, Sheraton J, Storms RK, Bussey H. Завершение 9Геном 0003 Saccharomyces cerevisiae позволяет идентифицировать KTR5 , KTR6 и KTR7 и определить девятичленное семейство генов KRE2/MNT1 маннозилтрансфераз в этом организме. Дрожжи. 1997; 13: 267–274. [PubMed] [Google Scholar]
36. Люссье М., Уайт А.М., Шератон Дж., Ди Паоло Т., Тредуэлл Дж., Саутхард С.Б., Хоренштейн С.И., Чен-Вайнер Дж. , Рам А.Ф., Каптейн Дж.С., Ремер Т.В., Во Д.Х., Бондок Д. К., Холл Дж., Чжун В. В., Сиду А. М., Дэвис Дж., Клис Ф. М., Роббинс П. В., Бусси Х. Крупномасштабная идентификация генов, участвующих в биосинтезе и архитектуре клеточной поверхности в Saccharomyces cerevisiae . Генетика. 1997; 147: 435–450. [Бесплатная статья PMC] [PubMed] [Google Scholar]
37. Маннерс Д. Дж., Массон А. Дж., Паттерсон Дж. К. Структура β1,3-D-глюкана из клеточных стенок дрожжей. Биохим Дж. 1973; 135:19–30. [PMC free article] [PubMed] [Google Scholar]
38. Montijn RC, Van Wolven P, De Hoog S, Klis FM. β-глюкозилированные белки в клеточной стенке черных дрожжей Exophiala (Wangiella) dermatitidis . Микробиология. 1997; 143:1673–1680. [PubMed] [Академия Google]
39. Mrsa V, Seidl T, Gentzsch M, Tanner W. Специфическое мечение белков клеточной стенки путем биотинилирования. Идентификация четырех ковалентно связанных O-маннозилированных белков Saccharomyces cerevisiae . Дрожжи. 1997; 13:1145–1154. [PubMed] [Google Scholar]
40. Muller A, Ensley H, McNamee R, Jones E, McLaughlin E, Chandley W, Browder W, Lowman D, Williams D. Применение различных протонных кислот для экстракции 1- 3β-D-глюкан из Saccharomyces cerevisiae . Карбогид Рез. 1997; 299: 203–208. [PubMed] [Google Scholar]
41. Muller G, Gross E, Wied S, Bandlow W. Индуцированный глюкозой последовательный процессинг гликозил-фосфатидилинозитол-заякоренного эктопротеина в Saccharomyces cerevisiae . Мол Селл Биол. 1996; 16: 442–456. [Бесплатная статья PMC] [PubMed] [Google Scholar]
42. Орлеан П. Биогенез дрожжевой стенки и поверхностных компонентов. В: Прингл Дж., Броуч Дж., Джонс Э., редакторы. Молекулярная и клеточная биология дрожжей Сахаромицеты . 3. Клеточный цикл и клеточная биология. Колд-Спринг-Харбор, Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор; 1997. С. 229–362. [Google Scholar]
43. Пополо Л., Джиларделли Д., Бонфанте П. , Вай М. Увеличение количества хитина как важный ответ на дефекты сборки полимеров клеточной стенки у мутанта ggp1 Δ Saccharomyces cerevisiae . J Бактериол. 1997; 179: 463–469. [Бесплатная статья PMC] [PubMed] [Google Scholar]
44. Ram A F J, Kapteyn J C, Montijn RC, Caro L H P, Douwes J E, Baginsky W, Mazur P, van den Ende H, Klis F M. Потеря плазмы мембраносвязанный белок Gas1p в Saccharomyces cerevisiae приводит к высвобождению β1,3-глюкана в среду и запускает механизм компенсации, обеспечивающий целостность клеточной стенки. J Бактериол. 1998; 180:1418–1424. [Статья бесплатно PMC] [PubMed] [Google Scholar]
45. Ram A F, Wolters A, Ten Hoopen R, Klis F M. Новый подход к выделению мутантов клеточной стенки в Saccharomyces cerevisiae путем скрининга на гиперчувствительность к кальцинированному белому . Дрожжи. 1994; 10:1019–1030. [PubMed] [Google Scholar]
46. Saito H, Yoshioka Y, Yoloi M, Yamada J. Различные механизмы гелеобразования между линейными и разветвленными 1-3β-D-глюканами, выявленные с помощью твердого состояния с высоким разрешением 13 С ЯМР. Биополимеры. 1990; 29: 1689–1698. [PubMed] [Google Scholar]
47. Schopfer P. Опосредованное перекисью водорода усиление клеточной стенки in vitro в колеоптилях кукурузы. Планта. 1996; 199:43–49. [Google Scholar]
48. Шоуолтер А. М. Структура и функция белков клеточной стенки растений. Растительная клетка. 1993; 5:9–23. [Статья бесплатно PMC] [PubMed] [Google Scholar]
49. Скшипек М., Лестер Р.Л., Диксон Р.С. Анализ генов-супрессоров показывает важную роль сфинголипидов в транспорте гликозилфосфатидилинозитол-заякоренных белков в Saccharomyces cerevisiae . J Бактериол. 1997; 179:1513–1520. [Бесплатная статья PMC] [PubMed] [Google Scholar]
50. Stokke B T, Elgsaeter A, Hara C, Kitamura S, Takeo K. Физико-химические свойства 1-6-разветвленных 1-3 β-D-глюканов. 1. Физические размеры, оцененные по гидродинамическим и электронно-микроскопическим данным. Биополимеры. 1993; 33: 561–573. [PubMed] [Google Scholar]
51. Табучи А., Камисака С., Хосон Т. Очистка ксилоглюкангидролазы/эндотрансферазы из клеточных стенок эпикотилей фасоли адзуки. Физиология клеток растений. 1997;38:653–658. [Google Scholar]
52. Van der Vaart JM, Caro LHP, Chapman JW, Klis FM, Verrips CT. Идентификация трех маннопротеинов в клеточной стенке Saccharomyces cerevisiae . J Бактериол. 1995;177:3104–3110. [Бесплатная статья PMC] [PubMed] [Google Scholar]
53. van Der Vaart JM, te Bisebeke R, Chapman JW, Klis FM, Verrips CT. β-1,6-глюкан, содержащий боковую цепь клеточной стенки белки Saccharomyces cerevisiae связаны с гликановым ядром фрагмента GPI. FEMS Microbiol Lett. 1996;145:401–407. [PubMed] [Google Scholar]
54. Фоссен Дж. Х., Мюллер В. Х., Липке П. Н., Клис Ф. М. Рестриктивный синтез якоря гликозилфосфатидилинозитола в дрожжевых клетках cwh6/gpi3 вызывает аномальный биогенез белков клеточной стенки. J Бактериол. 1997;179:2202–2209. [Бесплатная статья PMC] [PubMed] [Google Scholar]
55. Williams DL, McNamee RB, Jones EL, Pretus HA, Ensley HE, Browder IW, Di Luzio NR. Метод солюбилизации 1-3β-D -глюкан, выделенный из Saccharomyces cerevisiae . Карбогид Рез. 1991; 219: 203–213. [PubMed] [Google Scholar]
56. Yen PH, Ballou CE Частичная характеристика фактора половой агглютинации из клеток Hansenula wingei Y-2340 типа 5. Биохимия. 1974; 13: 2428–2437. [PubMed] [Google Scholar]
57. Zlotnik H, Fernandez P, Bowers B, Cabib E. Saccharomyces cerevisiae маннопротеины образуют внешний слой стенки, который определяет пористость. J Бактериол. 1984; 159: 1018–1026. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Клеточная стенка Определение и примеры
Клеточная стенка
сущ., множественное число: клеточные стенки
[sɛl wɑːl]
Определение: Структурный слой, окружающий клетку, расположенный рядом с клеточной мембраной
Содержание
Клетка структурная, функциональная и биологическая единица всех организмов. Это мембраносвязанная структура, содержащая цитоплазму и цитоплазматические структуры. Мембрана, окружающая клетку и отделяющая ее от внешней среды, называется клеточной мембраной, тогда как клеточная стенка представляет собой еще один структурный слой, окружающий клетку, рядом с клеточной мембраной.
Определение клеточной стенки
Что такое клеточная стенка и как она формируется? Можно просто определить клеточную стенку как полужесткую толстую защитную структуру, которая окружает клеточную мембрану некоторых типов клеток для защиты и определения формы клетки. Клеточная мембрана сама по себе не может обеспечить требуемую жесткость или прочность.
Все живые организмы состоят из клеток. Клетки отвечают за выполнение всех жизненно важных функций, таких как обмен веществ, размножение и выделение. Клетка состоит из внутриклеточных структур, таких как органеллы. Органеллы имеют специфические функции и встроены в цитоплазму. Клетка окружена плазматической мембраной (или клеточной мембраной), чтобы отделить содержимое клетки от внешней среды.
Биологическое определение: A клеточная стенка представляет собой структурный слой рядом с клеточной мембраной, основная роль которого заключается в придании клетке жесткости и защите от механических воздействий. Его основная функция заключается в придании клетке жесткости, прочности и защите от механических воздействий. Примерами организмов с клеточными стенками являются растения, грибы, протисты (особенно плесени и водоросли) и большинство бактерий (несколько исключений — микоплазма и L-форма бактерий) имеют клеточные стенки. Животные и гетеротрофные протисты не имеют клеточных стенок.
Вопрос: Все ли клетки имеют клеточную стенку?
Ответ: Не все клетки имеют клеточные стенки.
Вопрос: Итак, какие клетки имеют клеточную стенку?
Ответ: Прокариоты, за исключением микоплазмы и бактерий L-формы, и некоторые эукариоты имеют клеточную стенку.
Вопрос: Имеют ли клетки животных клеточную стенку?
Ответ: В клетках животных нет клеточных стенок. Вот почему клетки животных не такие жесткие, как другие клетки, окруженные клеточными стенками. Таким образом, клетки животных обладают большей гибкостью, чем клетки растений.
Клеточная стенка и клеточная мембрана
Клеточные стенки и клеточные мембраны различаются по своей структуре, функциям и другим характеристикам. Краткое изложение см. ниже.
| Таблица 1: Клеточная стенка в сравнении с клеточной мембраной и органеллы | |
|---|---|
| Толщина обычно от 0,1 мкм до нескольких мкм; но варьируется в зависимости от состава, т. е. если новая (вторичная) клеточная стенка откладывается старой (первичной) клеточной стенкой | Обычно толщина 7,5–10 нм |
| Компоненты различаются в зависимости от вида | Состав липидного двойного слоя, с углеводами и липопротеинами |
| Придает жесткость, придавая клетке более определенную форму | Имея только клеточную мембрану (отсутствует клеточная стенка), клетка более гибкая и может изменять форму по мере необходимости |
| Обеспечивает защиту, напр. против воздействия осмотического давления; более проницаем для малых молекул | Обеспечивает защиту, напр. будучи избирательным, регулирующим прохождение веществ, так что не все могут легко проникнуть в клетку, даже если они малы, из-за ее полупроницаемости; защита от осмотического давления только до определенной точки |
| Отсутствие рецепторов клеточной поверхности | Наличие рецепторов клеточной поверхности |
| Присутствует в растениях, грибах, простейших (например, водорослях и плесневых грибах) и бактериях | Присутствует во всех клетках |
Функции клеточной стенки
Что делает клеточная стенка? Каковы специфические биологические функции клеточной стенки? Основной функцией клеточной стенки является обеспечение защиты внутренних структур клетки, поскольку плазматическая мембрана считается хрупким слоем, который не может обеспечить аналогичную защиту от различных условий окружающей среды. Кроме того, клеточные стенки отвечают за обеспечение отличительных форм клеток. Когда внутреннее давление внутри клетки увеличивается из-за поступления воды, клеточная стенка препятствует расширению и разрыву клетки. Что делает клеточная стенка? Клеточная стенка контролирует прохождение молекул через клетку, пропуская только небольшие метаболические молекулы, тем самым защищая клетку от различных токсинов и лекарств. У многоклеточных организмов, имеющих клеточные стенки, они помогают склеиванию клеток, придавая им характерную форму.
У растений клеточная стенка отвечает за набухание растения. Клеточная стенка защищает растительную клетку от разрыва, когда в клетку попадает слишком много воды. Вместо того, чтобы лопнуть, клетка способна противостоять осмотическому давлению, оказываемому молекулами воды. Следовательно, клетка остается набухшей. Некоторые растительные клетки имеют клеточную стенку, состоящую из одного слоя. Другие растительные клетки имеют две: первичную и вторичную клеточные стенки. Второй клеточный слой содержит большое количество лигнина и, следовательно, помогает сделать клетку водонепроницаемой.
Рисунок 1: Статусы растительных клеток, основанные на чистом движении воды через клетку при помещении в разные растворы.
Основной функцией клеточной стенки является обеспечение жесткости и структурной поддержки . У растений придает набухание. Он удерживает растительную клетку от разрыва (осмотический лизис) при помещении в гипотонический раствор. Чрезмерный осмос предотвращается, когда осмотическое давление воздействует на клеточную стенку, помогая стабилизировать растительную клетку. Однако, если растительную клетку поместить в гипертоническую среду, клеточная стенка не может предотвратить потерю клеткой воды. Это приводит к сморщиванию клеток (или их вялости). В отношении используемого механизма читать ТУРГОРНОЕ ДАВЛЕНИЕ .
Структура клеточной стенки
Из чего состоит клеточная стенка? Строение клеточных стенок неодинаково у всех видов; это зависит от стадии развития и типа клеток. Первичная клеточная стенка растения состоит из пектина, целлюлозы и гемицеллюлозы, а также других закрепленных или встроенных полимеров, таких как суберин, лигнин или кутин. Клеточные стенки водорослей состоят из полисахаридов и гликопротеинов, таких как агар и каррагинан; эти молекулы отсутствуют в наземных растениях. Стенки бактериальных клеток состоят из пептидогликанов, тогда как клеточные стенки архей состоят из псевдопептидогликана, S-слоя или полисахаридов. Клеточные стенки грибов состоят из полимера хитина, N-ацетилглюкозамин .
Клеточные стенки растений
Имеют ли растительные клетки клеточные стенки? Да, растительные клетки имеют клеточную стенку, которая представляет собой структуру, покрывающую каждую растительную клетку. Клеточная мембрана сама по себе не может обеспечить структурную целостность растительных клеток. Следовательно, структура растительной клетки прочнее с клеточной стенкой. Однако именно по этой причине растительные клетки более жесткие и менее гибкие, чем клетки животных.
Части растительных клеток заключены в жесткую, растяжимую и тонкую первичную клеточную стенку во время их роста, состоящую из пектина , целлюлозы, и гемицеллюлозы . Однако основным компонентом клеточной стенки растений является целлюлоза. Что такое целлюлоза? Целлюлоза представляет собой полисахарид, состоящий из линейной цепи β-(1→4) связанных звеньев D-глюкозы: (C 6 H 10 O 5 ) n .
Клеточная стенка может откладывать под собой еще один слой клеточной стенки. Когда это происходит, старая клеточная стенка называется 9.0003 первичная клеточная стенка , а вновь отложенная клеточная стенка обозначается как вторичная клеточная стенка . Вторичная клеточная стенка представляет собой толстую и прочную клеточную стенку, расположенную между первичной клеточной стенкой и клеточной мембраной. Он богат лигнином. Что такое лигнин? Лигнин является наиболее распространенным веществом во вторичной клеточной стенке и состоит из фенольных соединений, образующих сложную сеть. Это делает клеточную стенку непроницаемой для воды.
Средняя пластинка находится между двумя первичными клеточными стенками . Это богатый пектином межклеточный материал, который склеивает соседние клетки. См. Рисунок 2.
Рисунок 2: клеточная стенка растения может состоять из одного или двух слоев. Первичная клеточная стенка отвечает за секрецию второго слоя, называемого вторичной клеточной стенкой, над плазматической мембраной.
Какова функция клеточной стенки растительной клетки? Функции клеточной стенки растений следующие:
- Supports cell structure and provides definite shape
- Encloses the cell
- Helps transport material across the plant cell and the environment and the transportation of fluids in channels
- Resists change in osmotic pressure
Из чего состоят стенки растительных клеток в различных типах растительных клеток? У высших растений полисахарид целлюлозы образует эластичные волокна. Они тесно связаны друг с другом гликановой поперечной связью. В первичных клеточных стенках целлюлоза представляет собой сеть, встроенную в пектин. Вторичные клеточные стенки содержат лигнин, который отвердевает и укрепляет клеточную стенку. Молекулы клеточной стенки связаны вместе, образуя сложную структуру.
Рисунок 3: Структура клеточной стенки растений. Источник: изменено Марией Викторией Гонзагой из BiologyOnline.com, из работ Molecular Expressions Cell Biology: Plant Cell Structure — Cell Wall и через BioRender.
От чего защищает растение клеточная стенка? Клеточная стенка растения защищает его от внешней среды и от разрыва . Дисбаланс концентрации растворенных веществ внутри и снаружи клетки создает давление, которое растягивает клеточную стенку наружу. Как выглядит растительная клетка из-за этого давления? Растительная клетка выглядит набухшей (набухшей).
youtube.com/embed/dXsXSIjR1G0?autoplay=1″ src=»data:text/html;https://www.youtube.com/embed/dXsXSIjR1G0?autoplay=1;base64,PGJvZHkgc3R5bGU9J3dpZHRoOjEwMCU7aGVpZ2h0OjEwMCU7bWFyZ2luOjA7cGFkZGluZzowO2JhY2tncm91bmQ6dXJsKGh0dHBzOi8vaW1nLnlvdXR1YmUuY29tL3ZpL2RYc1hTSWpSMUcwLzAuanBnKSBjZW50ZXIvMTAwJSBuby1yZXBlYXQnPjxzdHlsZT5ib2R5ey0tYnRuQmFja2dyb3VuZDpyZ2JhKDAsMCwwLC42NSk7fWJvZHk6aG92ZXJ7LS1idG5CYWNrZ3JvdW5kOnJnYmEoMCwwLDApO2N1cnNvcjpwb2ludGVyO30jcGxheUJ0bntkaXNwbGF5OmZsZXg7YWxpZ24taXRlbXM6Y2VudGVyO2p1c3RpZnktY29udGVudDpjZW50ZXI7Y2xlYXI6Ym90aDt3aWR0aDoxMDBweDtoZWlnaHQ6NzBweDtsaW5lLWhlaWdodDo3MHB4O2ZvbnQtc2l6ZTo0NXB4O2JhY2tncm91bmQ6dmFyKC0tYnRuQmFja2dyb3VuZCk7dGV4dC1hbGlnbjpjZW50ZXI7Y29sb3I6I2ZmZjtib3JkZXItcmFkaXVzOjE4cHg7dmVydGljYWwtYWxpZ246bWlkZGxlO3Bvc2l0aW9uOmFic29sdXRlO3RvcDo1MCU7bGVmdDo1MCU7bWFyZ2luLWxlZnQ6LTUwcHg7bWFyZ2luLXRvcDotMzVweH0jcGxheUFycm93e3dpZHRoOjA7aGVpZ2h0OjA7Ym9yZGVyLXRvcDoxNXB4IHNvbGlkIHRyYW5zcGFyZW50O2JvcmRlci1ib3R0b206MTVweCBzb2xpZCB0cmFuc3BhcmVudDtib3JkZXItbGVmdDoyNXB4IHNvbGlkICNmZmY7fTwvc3R5bGU+PGRpdiBpZD0ncGxheUJ0bic+PGRpdiBpZD0ncGxheUFycm93Jz48L2Rpdj48L2Rpdj48c2NyaXB0PmRvY3VtZW50LmJvZHkuYWRkRXZlbnRMaXN0ZW5lcignY2xpY2snLCBmdW5jdGlvbigpe3dpbmRvdy5wYXJlbnQucG9zdE1lc3NhZ2Uoe2FjdGlvbjogJ3BsYXlCdG5DbGlja2VkJ30sICcqJyk7fSk7PC9zY3JpcHQ+PC9ib2R5Pg==»>
Клеточная стенка растений: Клеточная стенка растительных клеток может состоять из двух слоев: (1) первичной клеточной стенки и (2) вторичной клеточной стенки . Первичная клеточная стенка обычно представляет собой тонкий, гибкий и растяжимый слой, состоящий из целлюлозы, пектина и гемицеллюлозы. Вторичная клеточная стенка представляет собой толстый слой, богатый лигнином, который укрепляет и делает водонепроницаемой стенку и формируется внутри первичной клеточной стенки, площадь поверхности которой перестала увеличиваться, когда клетка полностью выросла. Между основными стенами находится средняя пластинка , которая представляет собой межклеточный материал, богатый пектином, который склеивает соседние клетки вместе. Клеточная стенка очень важна для растений, поскольку она помогает противостоять осмотическому давлению.
Клеточные стенки грибов
Имеют ли грибы клеточные стенки? Грибы — это эукариоты, клетки которых содержат клеточные стенки. Клеточная стенка грибов защищает его от хищников. Грибковая клеточная стенка и клеточная мембрана соединены вместе. Они регулируют все отношения между внешней средой и клеткой. Многие сигнальные и синтетические пути способствуют образованию клеточных стенок.
Клеточная стенка гриба защищает внутренние клеточные структуры от давления и стресса. Он защищает клетку от разрыва, контролирует проницаемость клетки, поглощение молекул из внешней среды, определяет архитектуру клетки и обеспечивает жесткость. Помимо защитной функции, белки клеточной стенки могут опосредовать взаимодействие с внешней средой, так как обычно содержат рецепторы и адгезивные белки. Более того, клеточные стенки содержат антигенные белки, которые запускают иммунную систему хозяина во время инфекции для подавления роста и размножения патогенных грибков. Структура клеточной стенки состоит из различных слоев, где все компоненты, отложившиеся на самом внутреннем слое, контактируют с плазматической мембраной.
Клеточная стенка грибов в основном состоит из хитина, глюканов и гликопротеинов. Эти структуры отсутствуют в клетках человека и животных. Следовательно, противогрибковые препараты обычно нацелены на эти структуры, чтобы обеспечить селективность в отношении грибковых клеток, не затрагивая клетку-хозяина. Белки и полисахариды в клеточной стенке обычно связаны друг с другом, образуя гликопротеины.
Клеточная стенка грибов в основном состоит из полисахарида Glucans, который составляет примерно половину сухой массы компонентов клеточной стенки. Полимеры глюкана в основном состоят из 1-3 связанных звеньев глюкозы. Хитин является одним из сухих компонентов клеточной стенки грибов, он составляет около 2% сухих компонентов клеточной стенки дрожжей, в то время как у грибов он составляет около 10-20%. Грибы могут синтезировать хитин под действием фермента хитинсинтазы с использованием N-ацетилглюкозамина, затем он накапливается рядом с цитоплазматической мембраной во внеклеточном пространстве. Белки составляют около 30-50% сухой массы клеточной стенки дрожжей, в то время как они составляют около 20-30% клеточной стенки мицелиальных грибов. Белки клеточной стенки связаны с углеводами N- или O-связью, образуя гликопротеины.
Клеточные стенки грибов содержат меланин с высокой молекулярной массой и отрицательно заряженным пигментом. Меланин нерастворим в воде и защищает грибковую клетку от стрессоров, таких как ультрафиолетовый свет, высокая температура и токсины, чтобы она могла выжить внутри клетки-хозяина. Производство меланина является одним из факторов вирулентности грибов; это важно для распространения и инвазии клетки-хозяина. Более того, меланин может влиять на иммунный ответ хозяина и ингибировать фагоцитоз.
Рисунок 6: Схематическая диаграмма клеточной стенки грибов. Кредит: Майя и Рике — диаграмма, CC BY 3.0.
Клеточные стенки водорослей
Клеточные стенки водорослей структурно родственны клеточным стенкам растений. Они состоят из полисахаридов, таких как гликопротеины или целлюлоза. Некоторые красные и зеленые водоросли содержат микрофибриллы маннана, а клеточная стенка бурых водорослей содержит альгиновую кислоту. В клеточной стенке водорослей могут накапливаться различные вещества в виде ионов кальция и спорополленина. Диатомеи представляют собой группу водорослей, которые используют кремниевые кислоты для синтеза своих клеточных стенок. Синтез кремнезема экономит энергию клетки, так как требует меньше энергии во время синтеза.
Рисунок 5: Схематическая диаграмма клеточной стенки водорослей. Предоставлено: Скуэлья, Ф. (2019). Использование гидролитических ферментов для обработки биомассы водорослей. Текущая биотехнология, 5 (4), 296–304. Источник.
Рисунок 6: Схематическая диаграмма клеточной стенки бурых водорослей. Клеточная стенка бурых водорослей состоит в основном из структурно сложных полисахаридов. Авторы и права: Deniaud-Bouët, E., et al. (2017). Обзор клеточных стенок бурых водорослей и сульфатированных полисахаридов, содержащих фукозу: контекст клеточной стенки, биомедицинские свойства и ключевые исследовательские задачи. Углеводные полимеры, 175, 395–408. DOI.
Формы
Формы, напр. слизевики и водяные плесени — грибовидные протисты. Эти организмы сравнивают с грибами, потому что у них обоих отсутствуют хлоропласты, они гетеротрофны (сапрофиты) и имеют клеточные стенки. Разница между ними заключается в том, что у плесени клеточная стенка в основном состоит из целлюлозы и не содержит хитина. Клеточные стенки грибов, напротив, содержат хитин.
Рисунок 7: желтый слизевик на земле.
Протисты и клеточные стенки грибов: Клеточная стенка водорослей состоит из гликопротеинов и полисахаридов. Другие водоросли могут иметь кремниевую кислоту в своих клеточных стенках. Формы ( грибоподобные протисты ) также имеют клеточные стенки, и их клеточная стенка похожа на клеточную стенку растений тем, что состоит в основном из целлюлозы . Целлюлозная клеточная стенка, по сути, является одной из характеристик, отличающих плесневые грибы от грибов. В клеточной стенке грибов помимо других веществ, таких как полисахариды, есть хитин.
Клеточные стенки бактерий и архей
Большинство прокариот окружены клеточными стенками, которые защищают хрупкую цитоплазматическую мембрану и другие клеточные компоненты. Имеют ли бактерии клеточные стенки? Бактерии и археи — одноклеточные прокариоты. Клеточная стенка прокариот представляет собой полужесткую сложную структуру.
Прокариотическая клеточная стенка способствует патогенности некоторых видов и является местом действия нескольких противомикробных препаратов. Более того, клеточные стенки бактерий используются для дифференциации двух основных типов бактерий: грамположительных и грамотрицательных бактерий.
Стенка бактериальной клетки состоит из жесткой сети пептидогликана, которая может быть обнаружена либо в сочетании с другими молекулами, либо отдельно. Сеть пептидогликана состоит из повторяющихся звеньев дисахаридов, образующих решетку и связанных между собой полипептидами. Дисахаридные звенья состоят из моносахаридов, называемых N-ацетилмурамовой кислотой (NAM) и N-ацетилглюкозамином (NAG). Эти моносахариды структурно родственны глюкозе. Клеточная стенка пептидогликана состоит из рядов чередующихся единиц NAM и NAG, образующих основу из углеводов, связанных вместе полипептидами. Антимикробный препарат пенициллин повреждает клеточную стенку бактерий, нарушая связь между рядами пептидогликана, что приводит к лизису и разрыву клетки из-за потери ее целостности.
Биология определения грамположительной клеточной стенки представляет собой жесткий, толстый слой пептидогликана, образованный из нескольких рядов. Плазменное пространство находится между плазматической мембраной и клеточными стенками грамположительных бактерий. Это пространство содержит липотейхоевую кислоту, образующую зернистый слой. Кроме того, в клеточных стенках грамположительных бактерий обнаружены другие вещества, такие как фосфат и тейхоевая кислота, которые состоят в основном из спиртов, таких как рибит или глицерин. Тейхоевая кислота отвечает за регуляцию движения катионов через клеточную стенку благодаря своему отрицательному заряду. Кроме того, тейхоевая кислота предотвращает повреждение и лизис клеточных стенок, таким образом, она играет роль в клеточном росте. Кроме того, он придает клетке антигенный характер, поэтому ее можно легко обнаружить и протестировать.
Фигура 8: состав клеточной стенки грамположительных бактерий. Предоставлено: Twooars, CC BY 3.0.
Стенки клеток грамотрицательных бактерий состоят всего из нескольких слоев пептидогликана и внешней мембраны. Периплазматическое пространство наружной мембраны состоит из гелеобразного вещества, содержащего высокую концентрацию транспортных белков и ферментов деградации. Поскольку клеточные стенки грамотрицательных бактерий содержат всего несколько рядов пептидогликана и не содержат тейхоевой кислоты, следовательно, они более подвержены повреждению и разрушению внешними факторами.
Наружная мембрана состоит из липопротеинов, липополисахаридов и фосфатов. Наружная мембрана способствует патогенности грамотрицательных бактерий, так как избегает действия комплемента и фагоцитоза благодаря своему отрицательному заряду. Кроме того, он устойчив к солям желчных кислот, пищеварительным ферментам, тяжелым металлам, детергентам и некоторым антибиотикам. Однако внешняя мембрана содержит порины, которые представляют собой каналы, образованные из-за присутствия белков, которые обеспечивают некоторую проницаемость для проникновения метаболических молекул, таких как углеводы, аминокислоты и витамин B12.
Липополисахарид внешней оболочки содержит молекулу липида, известную как липид А. Липид А высвобождается из бактериальной клетки после смерти, он действует как эндотоксин, который вызывает симптомы бактериальной инфекции, такие как лихорадка, свертывание крови, кровеносные сосуды дилатация и шок. По сравнению с тейхоевой кислотой грамположительных бактериальных клеток молекула полисахарида O присоединена к липиду А, он обеспечивает специфическую антигенность, поэтому его используют для различения различных штаммов грамотрицательных бактерий.
Клеточные стенки архей представляют собой атипичные клеточные стенки, состоящие из белков и полисахаридов, но не из пептидогликана. Однако их клеточная стенка содержит молекулу, подобную пептидогликану, называемую псевдопептидогликаном или слоем псевдомуреина S. В нем отсутствует NAM и вместо него содержится N-ацетилталозаминуроновая кислота. При микроскопическом исследовании археи обычно кажутся похожими на грамотрицательные бактерии из-за отсутствия у них пептидогликана в структуре их клеточной стенки.
youtube.com/embed/JRHOjgwXBrI?autoplay=1″ src=»data:text/html;https://www.youtube.com/embed/JRHOjgwXBrI?autoplay=1;base64,PGJvZHkgc3R5bGU9J3dpZHRoOjEwMCU7aGVpZ2h0OjEwMCU7bWFyZ2luOjA7cGFkZGluZzowO2JhY2tncm91bmQ6dXJsKGh0dHBzOi8vaW1nLnlvdXR1YmUuY29tL3ZpL0pSSE9qZ3dYQnJJLzAuanBnKSBjZW50ZXIvMTAwJSBuby1yZXBlYXQnPjxzdHlsZT5ib2R5ey0tYnRuQmFja2dyb3VuZDpyZ2JhKDAsMCwwLC42NSk7fWJvZHk6aG92ZXJ7LS1idG5CYWNrZ3JvdW5kOnJnYmEoMCwwLDApO2N1cnNvcjpwb2ludGVyO30jcGxheUJ0bntkaXNwbGF5OmZsZXg7YWxpZ24taXRlbXM6Y2VudGVyO2p1c3RpZnktY29udGVudDpjZW50ZXI7Y2xlYXI6Ym90aDt3aWR0aDoxMDBweDtoZWlnaHQ6NzBweDtsaW5lLWhlaWdodDo3MHB4O2ZvbnQtc2l6ZTo0NXB4O2JhY2tncm91bmQ6dmFyKC0tYnRuQmFja2dyb3VuZCk7dGV4dC1hbGlnbjpjZW50ZXI7Y29sb3I6I2ZmZjtib3JkZXItcmFkaXVzOjE4cHg7dmVydGljYWwtYWxpZ246bWlkZGxlO3Bvc2l0aW9uOmFic29sdXRlO3RvcDo1MCU7bGVmdDo1MCU7bWFyZ2luLWxlZnQ6LTUwcHg7bWFyZ2luLXRvcDotMzVweH0jcGxheUFycm93e3dpZHRoOjA7aGVpZ2h0OjA7Ym9yZGVyLXRvcDoxNXB4IHNvbGlkIHRyYW5zcGFyZW50O2JvcmRlci1ib3R0b206MTVweCBzb2xpZCB0cmFuc3BhcmVudDtib3JkZXItbGVmdDoyNXB4IHNvbGlkICNmZmY7fTwvc3R5bGU+PGRpdiBpZD0ncGxheUJ0bic+PGRpdiBpZD0ncGxheUFycm93Jz48L2Rpdj48L2Rpdj48c2NyaXB0PmRvY3VtZW50LmJvZHkuYWRkRXZlbnRMaXN0ZW5lcignY2xpY2snLCBmdW5jdGlvbigpe3dpbmRvdy5wYXJlbnQucG9zdE1lc3NhZ2Uoe2FjdGlvbjogJ3BsYXlCdG5DbGlja2VkJ30sICcqJyk7fSk7PC9zY3JpcHQ+PC9ib2R5Pg==»>
Прокариотическая клеточная стенка: У бактерий клеточная стенка состоит из пептидогликана. Бактерии с клеточными стенками можно разделить на грамположительные и грамотрицательные. Грамположительные бактерии обладают толстой клеточной стенкой (из-за нескольких слоев пептидогликана и тейхоевых кислот), тогда как Грамотрицательные бактерии имеют относительно тонкую клеточную стенку (только с несколькими слоями пептидогликана). У архей клеточная стенка обычно лишена пептидогликана (за исключением группы метаногенов) и состоит из S-слоев гликопротеина, псевдопептидогликана или полисахаридов.
Попробуйте ответить на приведенный ниже тест, чтобы проверить, что вы уже узнали о клеточных стенках.
Викторина
Выберите лучший ответ.
1. Где расположена клеточная стенка?
Рассеяны в цитоплазме
Наружная часть клетки
Под клеточной мембраной
2.