



Генетика митохондрий у дрожжей. В клетках дрожжей не обнаружены митохондрии
Генетика митохондрий у дрожжей
Дрожжи занимают центральное место в исследованиях по генетике митохондрий, поскольку эти органеллы наиболее активны у таких организмов. Половой цикл одного из представителей сахаромицетов Saccharomycescerevisiaeможет быть изображён в виде такой схемы:
РИСУНОК
Представленная схема отражает существование гаплоидных и диплоидного клонов клеток с образованием гаплоидных аскоспор.
Существование внеядерной наследственности у дрожжей впервые было показано в работах Эфрусси с соавт., описавших мутацию petite. Мутантные клетки, выращенные на глюкозе, обладали пониженным дыханием и почти не содержали цитохромов а1,a3,b.
Генетичекий анализ показал, что мутация petiteимеет цитоплазматическую природу. Позднее было установлено, что митохондриальная ДНК цитоплазматических мутантовpetiteотличается по плавучей плотности и содержанию ГЦ-пар от ДНК митохондрий нормальных клеток этих дрожжей.
Позднее были получены цитоплазматические мутации устойчивости дрожжей к антибиотикам.
Расщепление и рекомбинация митохондриальных генов в зиготах послужили основой для построения генетических карт цитоплазматических систем.
Дальнейшие работы позволили обнаружить у дрожжей явление супрессивности(suppression– подавление), которое выражается в том, что при скрещивании дикой (нормальной) формы дрожжей с мутантной типаpetiteв зиготах происходит преимущественная репликация ДНК митохондрий мутантной формы. Через некоторое время аллель дикого типа не просто переходит в скрытое (рецессивное) или неактивное состояние, а исчезает совсем и уже никогда не проявляется у потомства.
Явление супрессивности можно рассматривать как одну из форм преимущественной передачи наследственности, при которой один родительский цитоплазматический геном берёт верх над другим. Этот процесс оказывается аналогичным материнскому наследованию у хламидомонады, о котором говорилось выше.
Позднее цитоплазматические гены были выявлены и у многих других грибов. Биохимические изменения в работе митохондрий у них также были связаны с нарушениями биосинтеза цитохромной системы.
Таким образом, описанные примеры позволили расширить арсенал экспериментальных методов для исследования цитоплазматической наследственности и показать высокую значимость цитоплазматической наследственности для обеспечения жизнедеятельности организмов.
Роль цитоплазматических генов в биогенезе клеточных органелл
С самого начала ХХ столетия цитологии постоянно подчёркивают генетическую непрерывность хлоропластов и митохондрий, а именно то, что эти органеллы возникают только из предшествующих органелл того же рода.
Изучение роли митохондриальных генов в биогенезе митохондрий показали, что там присутствуют:
специфические ферменты – РНК-полимеразы;
все три вида РНК: рибосомальные, транспортные, информационные;
особенные рибосомы.
Именно эти компоненты обеспечивают способность органелл к синтезу белка, управляемому их собственными полинуклеотидами. За счёт работы этой системы синтезируются белки только внутренней мембраны митохондрий. При этом почти все белки митохондриальных рибосом синтезируются в цитоплазме.
Изучение роли хлоропластных генов в биогенезе хлоропластов позволило выявить те же компоненты белок-синтезирующей системы, что и в митохондриях. Однако, спектр молекул белков, образующихся в хлоропластах, оказался значительно шире, что обусловлено, главным образом, значительно большей информационной ёмкостью ДНК хлоропластов. Позднее было установлено, что за счёт собственной ДНК в хлоропластах синтезируется большая субъединица центрального фермента ассимиляции СО 2– рибулозобисфосфат-карбоксилазы, фермента фосфорибулокиназы, некоторые белковые компоненты фотосистемыII, а также белки, участвующие в построении внутренней хлоропластной мембраны, в т.ч. мембран тилакоидов, и т.д.
studfiles.net
Митохондрии дрожжей - Справочник химика 21
Митохондрии присутствуют во всех клетках эукариот, использующих для дыхания кислород. Число митохондрий на клетку варьирует от 1 (у мельчайших трипаносом) до 3-10 (в некоторых ооцитах). Типичная клетка печени содержит более 1000 митохондрий [21]. Новые интересные данные получены при изучении митохондрий дрожжей [22]. Исследование серийных срезов через одну клетку (дополнение 1-В) показало, что все митохондрии связаны между собой. Таким образом, митохондрии дрожжевых клеток — это не отдельные органеллы, а единая сообщающаяся внутриклеточная структура. Насколько верно это для других-организмов — пока неясно. [c.34] Субъединичным состав цитохромоксидазы сильно варьирует в зависимости от источника выделения. Так, цитохром-с-оксидазы бактерий содержат 2—3 субъединицы, митохондрий дрожжей — [c.617] Два прерывистых гена в митохондриях дрожжей обладают исключительными генетическими свойствами, которые находятся в таком противоречии с традиционным представлением о гене, что могут получить объяснение только при исследовании ДНК на молекулярном уровне. Строение этих генов приведено на рис. 20.22. [c.258] Противоречит ли наличие интронов в геноме митохондрий дрожжей предположению о том, что митохондрии появились в результате возникшего в древности симбиоза, когда прототип бактерии был встроен в цитоплазму эукариотической клетки Нет, если предположить, что такое встраивание предшествовало потере интронов прокариотами, хотя при этом мы должны допустить независимую потерю способности к удалению интронов при эволюции бактерий и митохондрий позвоночных. [c.265]Хлоропласты имеют более сложный геном, чем митохондрии дрожжей и человека [39, 46] [c.63]
Цитохромоксидаза — мембранный белок, состоящий из ряда субъединиц. Интересно, что в процессе эволюции шло усложнение этого комплекса. Цитохромоксидазы различных бактерий содержат от 2 до 3 субъединиц, в митохондриях дрожжей их — 7— -8, а в митохондриях высших животных—12—13 субъединиц. [c.128]
Код, данный на рис. 3, является универсальным для белоксинтези-рующих систем бактерий и цитоплазмы всех эукариот, включая животных, грибы и высшие растения. Однако в живой природе имеются также и исключения. По крайней мере белоксинтезирующие системы митохондрий животных (млекопитающих) и грибов обнаруживают ряд отклонений от этого универсального кода. Так, в митохондриях изученных эукариотических организмов триптофан кодируется как UGG, так и UGA соответственно, UGA не является терминирующим кодоном. В митохондриях млекопитающих (человека) кодоны AGA и AGG — терминирующие и не кодируют аргинин. В митохондриях дрожжей вся кодоновая семья UU, U , UA и UG кодирует треонин, а не лейцин (хотя в митохондриях другого гриба, Neurospora, они кодируют лейцин, в соответствии с универсальным кодом). [c.16]
Особые РНК-полимеразы обеспечивают транскрипцию клеточных органелл эукариот — хлоропластов и митохондрий. В составе хлоропластной ДНК обнаружены гены, гомологичные генам, кодирующим а-, - и -субъединицы РНК-полимеразы Е. oli. Это, а также сходство нуклеотидной последовательности промоторов бактерий и хлоропластов свидетельствует о том, что РНК-полимераза хлоропластов должна быть сходна с РНК-полимеразой бактерий. РНК-полимеразы митохондрий состоят, по-видимому, всего из одной субъединицы, подобно РНК-полимеразам, кодируемым некоторыми бактериофагами, такими, как ТЗ и Т7. РНК-полимераза митохондрий дрожжей сходна с РНК-полнмеразами этих фагов по аминокислотной последовательности. Ген, кодирующий митохондриальную РНК-полимеразу, располагается в ядре. [c.136]
Отклонения от универсальности кода, как об этом уже говорилось на с, 262, наблюдаются в митохондриях. У митохондрий человека АУА кодирует не Иле, а Мет, кодон УГА — не обрыв цепи, а Трп, кодоны АГА и АГГ — терминирующие. Кодовая таблица оказывается более симметричной в соответствии с нред-скаэанием Ичаса (1969). У митохондрий дрожжей УГА—Трп ЦУА, ЦУЦ, ЦУГ, ЦУУ — не Лей, а Тре, АГА, АГТ — терминирующие. Вырождение для Тре достигло восьми кодонов. [c.279]
Митохондрии — это замкнутые клеточные полиморфные структуры с многочисленными перегородками, возникающие в результате постепенной инвагинации цитоплазматической мембраны. Размеры митохондрий варьируют в широких пределах. Форма митохондрий может быть удлиненной, эллипсовидной или круглой. Эти органоиды ответственны за энергетический обмен клетки и в зависимости от энергонапряженности обмена в клетке внутренняя мембрана может иметь меньше (не напряженный обмен) или больше (энергонапряженный обмен) складок или трубочек (крист). Наружная мембрана митохондрий дрожжей очень прочна и однородна. Внутренняя мембрана неоднородна, к ней в большом количестве прикреплены грибовидные структуры, которые, по-видимому, являются местом сосредоточения ферментов, вероятнее всего, участвующих в процессе окислительного фосфорилирования. Внутренняя мембрана митохондрий, особенно кристы, более лабильна, чем внешняя. [c.28]
Сходный тип супрессии был обнаружен в митохондриях дрожжей. При этом супрессируются мутации, вызванные вставками или делециями одного остатка Т на участке из пяти Т. [c.101]
Геном митохондрий дрожжей существенно больше. Его размер у различных штаммов S. erevisiae варьирует в широких пределах, составляя в среднем около 84 т. п. н. В дрожжевых клетках при стандартном определении обнаруживается около 22 митохондрий на одну клетку, что предполагает наличие примерно 4 копий генома в каждой органелле. В растущих клетках содержание митохондриальной ДНК может достигать 18% всей ДНК клетки. (Митохондриальные геномы других видов дрожжей могут иметь существенно большие размеры.) [c.282]
Данные о происхождении компонентов, необходимых для экспрессии генов в митохондриях дрожжей, приведены в табл. 22.3. В целом аналогичная картина наблюдается в митохондриях других видов организмов и в хлоропластах. В митохондриях дрожжевых клеток можно выделить восемь мРНК, и они транскрибируются с помощью РНК-полимеразы, которая (предположительно) кодируется ядерными генами. Только мРНК, транскрибируемые с митохондриальных генов, могут транслироваться внутри митохондрии, и, с другой стороны, это единственное место в клетке, где эти мРНК могут экспрессироваться. [c.283]
В табл. 22.4 приведены комплексы, содержащие белки, синтезированные в митохондриях дрожжей. АТРаза состоит из двух частей мембранного фактора, образуемого двумя или более субъединицами, кодируемых митохондриальным геномом, и растворимой АТРазы F1, состоящей примерно из пяти субъединиц, синтезируемых в цитоплазме. Цитохром-с—оксидаза также состоит из субъединиц, происходящих из обоих источников. В состав комплекса цитохромов Ьс входит один белок митохондриального происхождения, связанный с щестью субъеди- ницами цитоплазматического происхождения. Малая субъединица рибосомы включает в себя один белок (Уаг 1), кодируемый митохондриальными генами. Были получены мутации, позволяющие идентифицировать почти все митохондриальные гены. [c.284]
| Рис. 22.2. В состав генома митохондрий дрожжей входят имеющие прерывистое и непрерывное строение гены, кодирующие белки, гены, кодирующие рРНК, и гены, кодирующие тРНК (положение тРНК-генов не указано). Стрелки показывают направление транскрипции. |  |
Из нуклеотидных последовательностей границ сплайсинга и окружающих их участков очевидно, что Нй эхой комплементарности между соответствующими левой и прйюэй границами нет это исключает возможность их непосредственного соединения друг с другом путем спаривания оснований. Другая возможность состоит в специфическом попарном узнавании последовательностей на границах экзон—интрон белками или РНК. Модели такого рода могут быть применены к митохондриям дрожжей, где сплайсинг осуществляется только по нескольким парам границ (как описано в гл. 20). В случае гораздо большего числа границ в ядерных молекулах труднее проверить справедливость таких моделей. [c.325]
Аналогичные исследования генетического кода митохондриальных геномов дрожжей и Neurospora выявили определенные отличия как от универсального генетического кода, так и от кода, используемого в митохондриях человека. В митохондриях дрожжей и Neurospora, так же как и в человеческих митохондриях, триплет UGA считывается не как терминаторный, а как триптофановый кодон. В дрожжевом митохондриальном коде в отличие от всех остальных, включая универсальный, семейство триплетов типа UN кодирует не лейцин, а треонин. Трансляция всего этого семейства, так же как и в митохондриях других организмов, осуществляется с помощью одного типа молекул тРНК, несущих антикодон UAG. Как в дрожжевых, так и в человеческих [c.96]
О последовательностях нуклеотидов в митохондриальной ДНК высших растений сведений очень мало, но секвенирован почти полностью большой (78 ООО п. н.) митохондриальный геном дрожжей Sa haromy es erevisiae, и оказалось, что только около трети его кодирует белки. Эти данные говорят о том, что, возможно, большая часть избыточной ДНК в митохондриях дрожжей (а может быть, и высших растений) не имеет существенного значения для организма. [c.492]
Сходным образом осуществляется транспорт белков внутрь митохондрий. Если очищенные митохондрии дрожжей инкубировать с клеточным экстрактом, содержащим только что синтезированные радиоактивные дрожжевые белки, то можно наблюдать, что митохондриальные белки, кодируемые ядерным геномом, отделяются от немитохондриальных белков цитоплазмы [c.64]
ГО — ЛИШЬ В Присутствии дитионита. Кроме того, для цитохромов Ь этих двух типов были зарегистрированы различные спектры как в интактных митохондриях, так и в очищенном комплексе. Наконец, они имели различные потенциалы полувосстановления, и эта разница сохранялась даже в выделенных димерах цитохромов (von Jagow et al., 1978). Эти цитохромы были названы Ьк или S62 — цитохром, восстанавливаемый сукцинатом, и Ьт или Ьббб — цитохром, который восстанавливается лишь ди-тионитом. В дальнейшем, однако, генетические исследования на митохондриях дрожжей показали, что апобелки обоих цитохромов кодируются одним структурным геном и, следовательно, являются структурно идентичными субъединицами димера. В этом случае различное поведение цитохромов в димере, по-видимому, связано с взаимодействиями мономеров. [c.119]
Аналогичный прием был использован и в другой работе [Stephenson et al., 1980]. Ее авторы исследовали гидрофобные белки внутренней мембраны митохондрий дрожжей. В присутствии циклогексимида, блокирующего синтез белков в цитоплазме, белки митохондрий метили S, а затем экстрагировали и фракционировали в первом направлении методом ИЭФ в 4%-ном [c.51]
II про-мРНК митохондрий дрожжей также имеют консервативные последовательности, а возможно, и специфическую вторичную структуру, но она отличается от структуры интронов группы I. Интроны этого типа также способны к самовырезанию, но об особенностях их вторичной и третичной структуры и соответственно о молекулярных деталях сплайсинга известно меньше. Очевидны две особенности механизма сплайсинга 1) в отличие от самосплайсинга интронов группы I для сплайсинга интронов группы II не нужен нуклеозид-инициатор 2) в результате сплайсинга образуется структура типа лассо (рис. 8.76). [c.112]
Позднее было показано, что окисление лактата в митохондриях дрожжей идет в обход начальных и средних участков фосфорили-рующей дыхательной цепи. В обходном пути свободного окисления участвует цитохром Ьг, заключенный в межмембранном пространстве, а также цитохромоксидаза. НАД не нужен для окисления лактата дрожжевыми митохондриями. По данным А. Ф. Броди и сотрудников, клетки My oba terium phlei окисляют лактат быстрее, чем другие субстраты, но без фосфорилирования. [c.190]
chem21.info
МИТОХОНДРИИ ДРОЖЖЕЙ: ТОНКАЯ СТРУКТУРА
МИТОХОНДРИИ ДРОЖЖЕЙ: ТОНКАЯ СТРУКТУРА
Исследование ультратонких срезов митохондрий различных дрожжевых организмов [см. Бирюзова ea 1973 , Lloyd ea 1974 , Stevens ea 1977 , Stevens ea 1981 ] позволяет отметить принципиальное сходство их тонкой структуры с таковой высших организмов ( рис. 1 ). Снаружи дрожжевые митохондрии отграничены от цитоплазмы наружной двуслойной мембраной , оба слоя которой либо непосредственно соприкасаются друг с другом, либо разделены осмиофобным пространством. Изнутри наружную мембрану выстилает внутренняя мембрана . Она образует выступы - тубулы и или настоящие кристы, чаще всего пузырчато-трубчатого строения, достигающие 150-200 А в толщину и состоящие из двух осмиофильных слоев, разделенных пространством в 50-100 А. Кристы могут идти неравномерно во всех направлениях и не обязательно параллельно длинной оси митохондрий, как это сообщалось первыми исследователями. Внутреннее пространство между кристами заполнено матриксом, который, как правило, менее плотен, чем окружающая органоид цитоплазма. Новая информация о некоторых деталях строения митохондриальной структуры была получена при использовании недавно развитой техники негативного контрастирования и скалывания замороженных образцов (freeze- etching), обеспечивающих высокое разрешение и стабильность препаратов. Метод негативного контрастировапия позволяет воссоздать трехмерную пространственную структуру дрожжевых митохондрий. На микрофотографии ( рис. 2 ) отчетливо видна ограничивающая внешняя мембрана и находящиеся под ней складки внутренней мембраны, расходящиеся под разными углами. Кроме того, с помощью метода негативного контрастирования фосфорно- вольфрамовой кислотой была обнаружена новая субструктура дрожжевых митохондрий - регулярно расположенные мелкие грибовидные образования, покрывающие всю свободную поверхность внутренней мембраны и крист, обращенную в матрикс митохондрий (см. рис. 3 ). Детальное рассмотрение ультраструктуры частиц показывает, что каждая состоит из головки сферической формы с d=70-80 А, стебля длиной 45-50 А и шириной 20-40 А и базальной части, представляющей собой сегмент внутренней мембраны митохондрий. Аналогичные структуры найдены в митохондриях из других источников и, по- видимому, характерны для всех мембран, катализирующих окислительное фосфорилирование. Особенно плодотворным для изучения ультратонкой структуры дрожжевых митохондрий оказался метод фиксации клеток замораживанием [см. Фихше ea 1973 , Фихше ea 1973 ]. Модификации этого метода, основанные на сверхбыстром замораживании клеток с последующим нанесением реплик на обнаженные поверхности [см. Фихше ea 1973 , Фихше ea 1973 ], позволили выявить детали, не распознаваемые с помощью обычных методов фиксации. Так, с помощью этого метода показано, что митохондриальные мембраны содержат глобулярные частицы, упаковка которых носит уникальный характер. Слой внешней мембраны, прилегающий к цитоплазме, имеет характерную шероховатую поверхность и содержит перфорации ("поры") нерегулярного устройства, что предполагает возможность контакта между цитоплазмой и перимитохондриальным пространством. При протравливании препаратов парами платины на этой поверхности обнаруживаются многочисленные более мелкие, чем перфорации, частицы [см. Котельникова ea 1973 ]. Поверхности внешней и внутренней мембраны, разделенные незначительным пространством или примыкающие друг к другу, относительно гладкие и содержат лишь рыхло упакованные частицы. На поверхности внутренней мембраны, обращенной к матриксу, четко выявляются глобулярные, относительно плотно упакованные частицы, сопоставимые по своим размерам с теми, что видны при негативном контрастировании. Новые структурные элементы обнаружены и в матриксе. Они имеют форму непрерывной фиброзной сети с нерегулярными плотно упакованными гранулами и областями с концентрацией полос или борозд, представляющих собой, вероятно, также плотно упакованные частицы [см. Фихше ea 1973 , Фихше ea 1973 ].
Результаты структурного анализа хорошо коррелируют с биохимическими доказательствами локализации в матриксе высокополимерной ДНК и сложно организованных полиферментных комплексов и с данными о различном составе и функциональной нагрузке внешней и внутренней митохондриальных мембран (см. разд. 1.3 ). Как правило, митохондрии дрожжей содержат меньше крист и имеют тенденцию к нерегулярности в очертаниях, устройстве, форме и упаковке их, т. е. характеризуются менее упорядоченной и жесткой структурой, чем более сложные митохондрии высших организмов. Внутренняя митохондриальная мембрана варьирует от настоящих крист до тубулей и обнаруживает удивительную вариабельность в структурных деталях в зависимости от специфики обмена, условий культивирования и фазы роста дрожжевых клеток. Дрожжи с ярко выраженным аэробным типом обмена - облигатные аэробы и слабо бродящие факультативные анаэробы - отличаются развитым мембранным аппаратом и содержат большое число сложно структурированных митохондрий с многочисленными кристами. У бродящих видов дрожжей мембранный аппарат выражен слабо, имеются лишь немногочисленные крупные митохондрии с неправильно ориентированными кристами; у дрожжей низового брожения кристы рудиментируются. Однако следует подчеркнуть, что тонкая структура митохондрий не остается постоянной даже у одного и того же организма (особенно у факультативных анаэробов), а меняется в зависимости от физиологического состояния, фазы роста и условий культивирования. Поэтому перечисленные выше характерные морфологические особенности относятся лишь к вполне "сформировавшимся" митохондриям в клетках, собранных в поздней экспоненциальной или стационарной фазах роста. Те же клетки в экспоненциальной фазе роста или при выращивании на среде с высокой концентрацией глюкозы обладают немногочисленными митохондриями со сравнительно простой внутренней организацией. Нередко полиморфизм митохондрий можно наблюдать даже в пределах одной клетки. Еще большая вариабельность отмечена для формы и размеров митохондрий. И это неудивительно, так как функционирование митохондриального аппарата, как известно еще из прижизненного наблюдения с использованием цейтраферной съемки, происходит в условиях его непрерывной подвижности, сопровождающейся изменением размеров и форы. В процессе движения митохондрии могут собираться в крупные агрегаты или, наоборот, распадаться на более мелкие образования. Характерная ориентация и направленное движение митохондрий, иногда сопровождающиеся агрегацией в специфические формы, происходит при делении и почковании дрожжевых клеток.
Деление дрожжевых митохондрий начинается примерно к 1/5 клеточного цикла после начала ядерного деления и заканчивается за 1/5 до конца цикла [см. Котельникова ea 1973 ]. Такая регулярная периодичность предполагает существование тесной корреляции между ядерным и митохондриальным делениями. До недавнего времени полагали, что движение митохондрий, а также изменение конформации в зависимости от энергетического состояния пассивно регулируется содержанием ионов и воды в матриксе [ Ernster ea 1981 ].
Однако обнаружение актина в митохондриях [ Ethoh ea 1990 ] и подавление антагонистами кальмодулина , зависимого от дыхания набухания митохондрий, дает основание полагать, что некоторые функции подвижности митохондрий могут контролироваться эндогенными (собственными) Са2+ зависимыми системами.
Принято считать, что в дрожжах и дрожжеподобных организмах, собранных в стационарную фазу роста, если возможность глюкозной репрессии минимальна, число митохондрий на одну клетку постоянно и зависит от систематической принадлежности организма. Однако использование серийных ультратонких срезов с последующей реконструкцией хондриома (совокупности всех митохондриальных структур [ Suzuki ea 1976 ]) показало, что в ряде случаев многочисленные отдельные митохондриальные профили, выявляемые на одиночных срезах, представляли собой поперечные сечения одной (или нескольких) ветвящейся трубчатой гигантской митохондрии. Так, в дрожжах Saccharomyces cerevisiae была обнаружена гигантская митохондрия длиной 50-60 мкм и диаметром 0,2-0,6 мкм [ Hoffman ea 1973 ]. Гигантские митохондрии найдены и в Schizosaccharomyces pombe [ Davison ea 1975 , Davison ea 1977 ], Candida utilis [ Davison ea 1975 , Davison ea 1977 , Keyhani ea 1980 ] и других видах дрожжей и дрожжеподобных организмов [см. Keddie ea 1969 ] и, следовательно, число митохондрий на одну клетку, по-видимому, меньше, чем полагали раньше. По данным [ Suzuki ea 1976 ], число митохондрий в клетке зависит от фазы роста и стадии клеточного цикла . Как правило, в экспоненциально растущих клетках число митохондрий меньше, чем в клетках, собранных в стационарной фазе роста. Предполагается, что такой циклический процесс фрагментации и слияния митохондрий облегчает рекомбинацию митохондриальных генов [ Grimes ea 1974 ].
Ссылки:
medbiol.ru
Ученые впервые локализовали все белки в митохондрии дрожжей
Немецкие исследователи впервые раскрыли локализацию каждого белка в митохондрии. Данное исследование поможет расшифровать метаболические пути и понять механизм многих заболеваний. Результаты работы опубликованы в Nature Communications.
Митохондрии — это электростанции клетки. Они преобразуют энергию из питательных веществ для любых целей клетки. Нарушение работы митохондрий может быть причиной заболеваний, особенно органов с быстрым метаболизмом, таких как мозг и сердце. Митохондрии состоят из четырех компонентов: одна внешняя и одна внутренняя мембрана, межмембранное пространство и матрица, которая является самой внутренней реакционной камерой. Каждый из этих компонентов имеет собственный белковый состав для выполнения конкретных функций. В дополнение к обеспечению энергией митохондрии выполняют другие важные метаболические задачи, например, в ней содержатся белки, контролирующие запрограммированную смерть клеток. Существует около 1500 различных видов белков в митохондрии у людей, в то время как S. cerevisiae (пекарские дрожжи) - популярная модель у ученых, имеют 1000. До сих пор локализация белков внутри органеллы остается плохо определенной и неточной. Исследование локализации и окружение белка (ландшафт) является важным для понимания точного механизма многих метаболических путей, а также определения возможных новых функций ранее неизвестных белков.
Для получения карты ландшафта митохондриальных белков у S. cerevisiae использовали подход, объединяющий стабильную изотопную маркировку, различные стратегии обогащения и экстракции белков и количественную масс-спектрометрию.
Дизайн эксперимента
F.-Nora Vögtle et al., 2017 / Nature Communications
Этот подход позволил классифицировать 986 белков и локализовать 818 белков, то есть ученые смогли получить точную карту нахождения белков в митохондрии. Также исследователи идентифицировали 206 белков, которые ранее не были аннотированы как митохондриальные. Таким образом, данное исследование может служить основой для изучения потенциальных новых функций митохондрий и лучшего понимания не только центральных биохимических процессов в клетках, но и развития многих заболеваний.
Раннее мы писали о роли митохондрий в патогенезе некоторых заболеваний. Например, была установлена связь между нарушением функций митохондрий и развитием расстройств аутического спектра.
Лилия Фахранурова
nplus1.ru
МИТОХОНДРИИ ДРОЖЖЕЙ: ИСТОРИЯ ОТКРЫТИЯ
МИТОХОНДРИИ ДРОЖЖЕЙ: ИСТОРИЯ ОТКРЫТИЯ
Впервые присутствие в цитоплазме дрожжевых клеток слабо преломляющих свет образований, способных окрашиваться цитохимическими красителями так же, как и митохондрии высших организмов, было отмечено в 1913 г.[см. Котельникова ea 1973 и ссылки в ней]. Первоначально они были выявлены на фиксированных и окрашенных препаратах, позднее были разработаны методы их идентификации в световом, фазово-контрастном и люминесцентном микроскопах, найдены специфические прижизненные красители, способные избирательно накапливаться, окрашивая эти органеллы, при сохранении полной жизнеспособности клеток. Стало ясно, что эти органеллы, описываемые как хондриосомы , митохондрии или хондриоконты , в зависимости от того, имели ли они форму нитей, палочек или зерен, являются универсальными элементами микроскопической структуры цитоплазмы дрожжевых клеток, их почек и спор. Более того, методы прижизненного наблюдения, позволяющие судить о поведении всей клеточной популяции, привели к установлению важных закономерностей: было определено число митохондрий на одну дрожжевую клетку; их локализация в клетке; структурно-функциональные отношения с другими компонентами клетки; обнаружена активная подвижность митохондрий с концентрированием их в зонах наиболее интенсивного роста и функционирования; выявлены их пластичность, полиморфизм, высокая сорбционная активность и способность к характерным перестройкам при изменении культивирования (детальное обсуждение вопроса и сводку литературы до 1972 г. см [ Котельникова ea 1973 ]). Таким образом, уже к 50-м годам в клетках дрожжей цитологически, цитохимически и функционально были идентифицированы структуры, сходные с митохондриями высших организмов. Данные, полученные с помощью метода электронной микроскопии, подтвердили сделанное ранее заключение об обязательном присутствии митохондриальных структур в дрожжевых клетках, находящихся на разных стадиях развития и размножения, почках и спорах.
Ссылки:
medbiol.ru
Митохондриальный геном дрожжей - Справочник химика 21
Митохондриальный геном дрожжей имеет большие размеры [c.284]Все классы генов могут иметь прерывистое строение все гены, кодирующие белки, а также гены, кодирующие рРНК, и гены, кодирующие тРНК. Интроны обнаружены также в митохондриальных генах дрожжей и хлоропластных генах. Прерывистые гены, по-видимому, присутствуют в клетках эукариот всех классов, хотя их содержание варьирует. Например, их доля среди ядерных структурных генов позвоночных может превышать таковую у грибов. [c.253]
Соответствие по крайней мере некоторых экзонов белковым доменам подтверждает предположение о том, что оно имеет фундаментальное значение в эволюции генов. Ясно, что дупликации и слияние экзонов могли играть важную роль в эволюции. Мы не можем проследить за действительными событиями, проишедшими в ходе эволюции каждого гена. Имеется несколько примеров взаимоотношений между экзонами и белковыми доменами, когда отсутствует их простое соответствие, но это можно объяснить тем, что такие события, как слияние экзонов, изменили структуру гена-предка в процессе эволюции ядерных генов. Однако в ряде случаев мы сталкиваемся с большими несоответствиями между структурами генов и белков. Митохондриальные гены дрожжей и млекопитающих кодируют практически идентичные митохондриальные белки, несмотря на существенные различия в организации генов. Геном митохондрий позвоночных очень мал и имеет чрезвычайно компактную организацию нерасщенленных генов (гл. 22), тогда как митохондриальный геном дрожжей имеет большие размеры и включает ряд сложных прерывистых генов. Какая форма гена была исходной [c.265]О последовательностях нуклеотидов в митохондриальной ДНК высших растений сведений очень мало, но секвенирован почти полностью большой (78 ООО п. н.) митохондриальный геном дрожжей Sa haromy es erevisiae, и оказалось, что только около трети его кодирует белки. Эти данные говорят о том, что, возможно, большая часть избыточной ДНК в митохондриях дрожжей (а может быть, и высших растений) не имеет существенного значения для организма. [c.492]
У дрожжей интроны могут иметься в митохондриальном гене одного штамма, но отсутствовать в том же гене другого штамма. Но-видимому, такие факультативные интроны способны включаться в гены и выходить из них подобно транспозонам. С другой стороны, в некоторых митохондриальных генах дрожжей интроны занимают те же позиции, что и в митохондриях Aspergillus и Neurospora значит, они были унаследованы от общего предка этих трех грибов. Вероятно, интроны имеют древнее происхождение, и хотя они были утрачены многими бактериями, они сохранились в геномах тех органелл, где регуляция сплайсинга РНК помогает контролировать экспрессию гена (разд. 10.5.5). [c.493]
Открытие интронов в митохондриальных генах дрожжей (а также и в некоторых генах хлоропластов) было неожиданным с точки зрения эндосимбио-тической теории происхождения митохондрий, так как в геноме бактерий интроны не обнаружены. Кроме того, интроны часто имеются в митохондриальном гене одного штамма дрожжей, но отсутствуют в том же гене у другого штамма. По этой и другим причинам было высказано предположение, что интроны в ДНК некоторых биоэнергетических органелл представляют собой остатки каких-то перемещающихся элементов (разд. 8.6. 0. Однако до сих пор не выяснено ни происхождение этих расщепленных генов в ДНК органелл, ни их влияние на организм. [c.64]
Особые типы интронов группа II. Интроны группы II распространены менее широко, чем интроны группы I. Они обнаружены в двух митохондриальных генах дрожжей, кодирующих одну из субъединиц цитохромоксвдазы и цитохром Ь интересно, что в этих генах присутствуют также интроны группы I. Интроны группы II не содержат канонических [c.108]
Было обнаружено существование интересного сходства между митохондриальными интронами дрожжей и некоторыми генами рРНК. В их состав входит несколько довольно коротких сходных последовательностей. Они расположены на некотором расстоянии от границ экзон—интрон (на самих границах таких консервативных последовательностей нет). Некоторые последовательности причастны к сплайсингу, по крайней мере, в случае гена box, поскольку они обеспечивают создание сайтов, соответствующих 1/ш -мутациям box 9 и box 2, блокирующим сплайсинг. [c.260]
По-видимому, все митохондриальные интроны дрожжей имеют такие канонические последовательности. Так, их обнаруживают в интронах, имеющих открытые кодирующие участки, и в других интронах, в которых все возможные рамки считывания заблокированы. К ядерным генам, содержащим канонические последовательности, относится ген Tetrahymena, кодирующий предшественник рРНК, подвергающийся автономному сплайсингу. Эти данные служат замечательной демонстрацией существования неожиданного эволюционного сходства ядерного и внеядерных геномов и позволяют сделать предположение о том, что, возможно, сплайсинг разных РНК протекает в соответствии с общим механизмов. Это заключение наводит на мысль об интересном парадоксе, касающемся механизмов сплайсинга. [c.261]
Данные о происхождении компонентов, необходимых для экспрессии генов в митохондриях дрожжей, приведены в табл. 22.3. В целом аналогичная картина наблюдается в митохондриях других видов организмов и в хлоропластах. В митохондриях дрожжевых клеток можно выделить восемь мРНК, и они транскрибируются с помощью РНК-полимеразы, которая (предположительно) кодируется ядерными генами. Только мРНК, транскрибируемые с митохондриальных генов, могут транслироваться внутри митохондрии, и, с другой стороны, это единственное место в клетке, где эти мРНК могут экспрессироваться. [c.283]
В табл. 22.4 приведены комплексы, содержащие белки, синтезированные в митохондриях дрожжей. АТРаза состоит из двух частей мембранного фактора, образуемого двумя или более субъединицами, кодируемых митохондриальным геномом, и растворимой АТРазы F1, состоящей примерно из пяти субъединиц, синтезируемых в цитоплазме. Цитохром-с—оксидаза также состоит из субъединиц, происходящих из обоих источников. В состав комплекса цитохромов Ьс входит один белок митохондриального происхождения, связанный с щестью субъеди- ницами цитоплазматического происхождения. Малая субъединица рибосомы включает в себя один белок (Уаг 1), кодируемый митохондриальными генами. Были получены мутации, позволяющие идентифицировать почти все митохондриальные гены. [c.284]
На рис. 22.2 приведена карта митохондриального генома дрожжей, показывающая взаимное расположение генов для основной части РНК и белков (за исключением молекул тРНК, число видов которьа составляет около 22 и гены для которых до сих пор полностью не картированы). Наиболее характерная особенность этой карты — пространственная разделенность локусов. [c.285]
Отличительная особенность митохондриального генома дрожжей состоит в пространственной разобщенности генов, кодирующих рРНК, что встречается крайне редко. Ген, кодирующий 15S-pPHK, непрерывен и располагается на расстоянии около 25 000 п.н. от гена для 21S-pPHK. В некоторых штаммах дрожжей последний ген имеет один интрон (как показано на карте), в других штаммах он не прерывается. [c.285]
Весьма значительная часть митохондриального генома дрожжей (около 25%) состоит из коротких (А—Т)-бо-гатых участков ДНК, вероятно не несущих функции кодирования. Однако значительная часть генетического материала все еще не изучена, и будет удивительно, если для оставшихся незаполненными участков карты не будут обнаружены другие гены. Но, даже допуская такую возможность, можно все же считать, что общее число генов в митохондриях дрожжей вряд ли превышает 20. [c.285]
В отличие от человека у некоторых растений и грибов (включая дрожжи) митохондриальные гены содержат интроны, которые должны быть удалены из транскрипта с последующим сплайсингом (разд. 3.2.7). У растений интроны обнаружены также примерно в 20 генах хлоропластов. Многие интроны в генах органелл содержат родственные нуклеотидные последовательности, которые могут исключаться из РНК-транскриптов в результате реакции, катализируемой самой РНК (разд. 9.4.14). хотя в этом самосплайсинге обычно участвуют и белки. Открытие интронов в генах органелл было неожиданным с точки зрения эндосимбиотической теории происхождения энергопреобразующих органелл, гак как в генах бактерий, от предков которых могли произойти митохондрии и хлоропласты, интронов не обнаружено. [c.493]
Но ряду причин большинство экспериментов по изучению механизмов биогенеза митохондрий проводится на культурах Sa haromy es arlshergensis (пивные дрожжи) и S. erevisiae ( пекарские дрожжи). Во-первых, при росте на глюкозе эти дрожжи обнаруживают уникальную способность существовать только за счет гликолиза и поэтому могут обходиться без функционально активных митохондрий, т.е. без окислительного фосфорилирования. Это дает возможность работать с клетками, митохондриальная и ядерная ДНК которых несут мутации, препятствующие нормальному развитию митохондрий. Такие мутации летальны почти у всех организмов. Во-вторых, дрожжи - простые одноклеточные эукариоты - легко выращивать и подвергать биохимическим исследованиям. И наконец, у дрожжей, обычно размножающихся бесполым способом путем почкования (асимметричного митоза), встречается и половой процесс. При половом размножении две гаплоидные клетки сливаются, образуя диплоидную зиготу, которая затем либо делится путем митоза, либо претерпевает мейоз и снова дает гаплоидные клетки. Возможность контролировать в лабораторных условиях чередование бесполого и полового размножения (разд. 13.2) намного облегчает проведение генетического анализа. Такой анализ позволяет выявить гены, ответственные за функцию митохондрий, и установить, которые из них находятся в ядерной ДНК и которые - в митохондриальной, поскольку мутации митохондриальных генов не наследуются по законам Менделя, которым подчиняется наследование ядерных генов [c.493]
Количество ДНК в некоторых низших эукариотах (дрожжи, Меиговрога) гораздо выше, чем в митохондриях высших животных. Это можно объяснить утратой генов и передачей их функций ядерному геному в ходе эволюции от низших эукариотов к высшим. С эволюционной точки зрения такое уменьшение может быть даже необходимым 1335]. Вероятно, оставшиеся гены представляют собой некоторый минимум и дальнейшая утрата генов по каким-то неизвестным нам причинам невозможна, а быть может, митохондриальные гены целиком находятся на пути к выходу 250]. [c.186]Митохондриальная транслирующая система тоже имеет общие черты с бактериальными белоксинтезирующими системами рибосомы митохондрий чувствительны к антибактериальным антибиотикам, синтез белка начинается с N-формилметионина. Однако есть и существенные различия. Самые поразительные из них выявляются при сопоставлении нуклеотидных последовательностей митохондриальных генов с аминокислотной последовательностью кодируемых ими белков. Например, триплет UGA, который служит в универсальном генетическом коде терминирующим кодоном, в митохондриях млекопитающих и дрожжей кодирует триптофан. Кроме того, отличаются значения нескольких других кодонов, причем здесь есть даже различия между кодами, действующими в митохондриях млекопитающих и дрожжей (табл. 9-4). Эти последние различия обусловлены особенностями митохондриальных тРНК, которые кодируются митохондриальным геномом и свойства которых мы подробнее рассмотрим позже. Почему генетический код в митохондриях отличен от кода бактерий и эукариот, пока не ясно. [c.58]
| Рис. 9-65. Здесь схематически показано различие в наследовании митохондриальных и ядерных генов у дрожжей. Две дрожжевые клетки из четырех, образовавшихся в результате мейоза, получают тот или иной ядерный ген от одного родителя, а две другие-от другого (менделевское наследование). В отличие от этого при передаче митохондриальных генов легко может случиться, что в период вегетативного роста (см. текст) все четыре клетки, образовавшиеся в результате мейоза, получат эти гены от одного и того же родителя (неменделевское, или цитоплазматическое, наследование). | 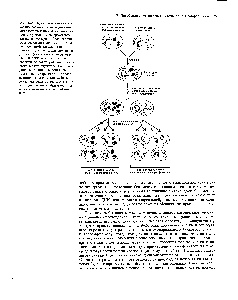 |
Последствия цитоплазматической передачи генов для некоторых животных, в том числе и для человека, более серьезны, нежели для дрожжей. Две сливающиеся гаплоидные дрожжевые клетки 1шеют одинаковую величину и вносят в образующуюся зиготу одинаковое количество митохондриальной ДНК. Таким образом, у дрожжей митохондри шьный геном наследуется от обоих родителей, которые вносят равный вклад в генофонд потомства (хотя, как мы уже видели, спустя несколько генераций отдельные потомки нередко будут содержать митохондрии только одного из родительских типов). В отличие от этого у высших животных яйцеклетка вносит в зиготу больше цитоплазмы, чем спермий, а у некоторых животных спермин могут вообще не вносить цитоплазмы. Поэтому можно думать, что у высших животных митохондриальный геном будет передаваться только от одного родителя (а именно по материнской линии) и действительно, это было подтверждено экспериментами. Оказалось, например, что при скрещивании крыс двух лабораторных линий с митохондриальной ДНК, слегка различающейся по последовательности нуклеотидов (типы А и В), получается потомство, содержащее митохондриальную ДНК только материнского типа. [c.60]
В митохондриальном геноме дрожжей с помощью генетических методов в сочетании с другими подходами были обнаружены два гена рРНК, примерно 30 генов тРНК и 8 генов, кодирующих белки. Семь из этих восьми белков локализованы во внутренней митохондриальной мембране, а один входит в состав рибосом. Все эти белки являются составными частями больших ферментных комплексов, образованных в основном из субъединиц, импортируемых органеллой из цитоплазмы. [c.61]
У дрожжей для кодирования всех известных продуктов митохондриальных генов нужно менее 20% информации, которая может быть потенциально заключена в ДНК митохондрий, но все попытки найти в ней какие-то дополнительные гены не привели к успеху. Причина такого противоречия-то, что митохондриальная ДНК дрожжей содержит очень много некодирующих участков, которые можно разделить на три категории. Во-первых, в некоторых генах имеются длинные вставочные последовательности (интроны) например, гены цитохрома Ь и субъединицы I цитохромоксидазы почти в десять раз больше, чем можно было бы ожидать, исходя из размера кодируемых ими белков. Разные штаммы могут резко отличаться друг от друга по числу и длине таких интронов. Во-вторых, некоторые митохондриальные мРНК имеют некодирующие лидерные последовательности, которые намного длиннее кодирующих участков. В-третьих, около половины митохондриальной ДНК дрожжей состоит из сегментов, на 95% образо- [c.61]
Генетические методы, столь успешно использованные при изучении митохондриального генома дрожжей, неприменимы в случае клеток человека. Однако благодаря своим относительно малым размерам наша митохондриальная ДНК-весьма подходящий объект для определения нуклеотидных последовательностей с помощью современной методики (разд. 4.5.4), и в 1981 г. была опубликована полная первичная структура молекулы этой ДНК, содержащей 16569 нуклеотидов. Сопоставляя эту структуру с уже известными нуклеотидными последовательностями митохондриальных тРНК и аминокислотными последовательностями белков, кодируемых митохондриальной ДНК, удалось определить локализацию многих генов в кольцевой молекуле ДНК из митохондрий человека (рис. 9-67). [c.62]
У некоторых дрожжей, в отличие от человека, митохондриальные гены содержат интроны, которые, вероятно, удаляются из мРНК с последующим сплайсингом. Однако нет никаких данных о симметричной транскрипции у дрожжей все гены, кроме одного, транскрибируются с одной цепи ДНК, причем существует несколько промоторов для различных генов. [c.64]
Некоторые особенности митохондриальной ДНК дрожжей (S. erevisiae). Гены, кодирующие белки, окрашены более интенсивно, а гены, кодирующие РНК,- светлее. Вставочные последовательности выделены точками (некоторые из них содержат открытые рамки считывания часть их указана). Серым цветом выделены неохарак-теризованные области некоторые из них содержат сайты начала репликации. Транскрипция в основном идет по часовой стрелке единственное известное исключение-транскрипция генов тРНК, начинающаяся в районе двух часов (отмечена стрелкой). [c.220]
Хлоропластные интроны, по-видимому, аналогичны или интронам группы I, или интронам группы II (разд. 8.5. в). Можно предположить, что при сплайсинге используются сходные механизмы. В большинстве хлоропластных интронов протяженные открытые рамки считывания отсутствуют. Впрочем, такая рамка длиной 509 кодонов обнаружена в интроне гена лизиновой тРНК в хлоропластах табака и в интроне гена 23S-pPHK у С. reinhardii. В соответствующих белковых продуктах имеются участки, гомологичные участкам матураз, кодируемым митохондриальными интронами дрожжей. Таким образом, интроны как хлоропластных, так и митохондриальных ДНК могут содержать функциональные сегменты. [c.225]
chem21.info
Митохондриальная ДНК дрожжей - Справочник химика 21
В случае митохондриальных интронов дрожжей белки не только специфичны по отношению к индивидуальным интронам, но и кодируются интронами, на которые затем и воздействуют. (Эта модель не применима ко всем ми- [c.261]Является ли специфическое разделение труда, демонстрируемое митохондриями и ядрами дрожжей, характерным и для других видов организмов Общее строение агрегатов, состоящих из белков со смешанным происхождением, сходно по крайней мере у митохондрий других грибов и млекопитающих. В геноме митохондрий дрожжей и млекопитающих присутствуют гены, кодирующие одни и те же белки. Таким образом можно сделать общее предположение о наличии значительной консервативности кодирующих функций митохондриальных геномов у различных видов организмов. [c.284]
Митохондриальный геном дрожжей имеет большие размеры [c.284]Перестройки митохондриальной ДНК дрожжей [c.288]
Общий вывод, который можно сделать на основе изучения митохондриальной ДНК петит-штаммов и штаммов дикого типа,-это значительная гибкость организации генома в отношении процессов его экспрессии и репликации. Возможно, что ее отличие от других митохондриальных ДНК обусловливает способность дрожжей существовать при полном отсутствии функционирования митохондрий. Было бы интересно выяснить, какие ферментные системы работают на этой ДНК. [c.288]
Подавляющее количество ДНК сосредоточено в ядре, обычно лишь небольшая часть ДНК находится в составе генома цитоплазматических органелл. Митохондрии грибов и млекопитающих содержат менее 1 % всей ДНК, а пластиды растений — 1—10 %. В клетках дрожжей Sa haromy es erevisiae количество митохондриальной ДНК может достигать 20 "6 от всей клеточной. [c.186]
В р-ции 1 цикла, катализируемой цитрат-оксалоацетат-лиазой, СНзС(0)8КоА стереоспецифично конденсируется с карбонильной группой оксалоацетата с образованием цитрата и свободного KoASH. Р-ция сопровождается значит, изменением своб. энергии (ДС — 32,24 кДж/моль) и является практически необратимой. Активность митохондриального фермента у дрожжей ингибируется АТФ. [c.634]
Цитохромоксидазы выполняют в аэробных организмах уникальную функцию они соединяются с Ог почти таким же образом, как и гемоглобин, а затем быстро восстанавливают Ог до двух молекул НгО [24а]. Происходит разрыв связи О—О для восстановления требуется четыре электрона. Очевидно, процесс этот сложен и пока еще плохо изучен. Важно отметить, что цитохромоксидаза, содержащаяся в митохондриях млекопитающих, имеет два гема (цитохром а) и два атома u(I) на одну функциональную единицу. Таким образом, при восстановлении обеих молекул цитохрома а и двух атомов меди может быть запасено четыре электрона для последующего восстановления одной молекулы Ог. Химия цитохромоксидазы слабо изучена. Как впервые обнаружил Кейлин, только половина молекул цитохрома а соединяется с СО. Она была названа цитохромом аз. По данным электрофореза в полиакриламидном геле с додецилсульфатом натрия, в цитохромоксида-зе дрожжей имеется шесть или семь субъединиц с мол. весом от 5 000 до 42 000 [24Ь, с]. Интересно отметить, что три наиболее крупные субъединицы, по-видимому, кодируются генами митохондриальной ДНК. Группы гема присоединены к пептидам меньшего размера. Было высказано предположение, что в интактном ферменте молекула Ог вначале связывается между атомом железа цитохрома аз и ионом двухвалентной меди aV—Ог—Си+. На следующей стадии происходит двухэлектронный процесс восстановления Ог с образованием перекисной структуры и далее двух молекул воды. [c.376]
Особые РНК-полимеразы обеспечивают транскрипцию клеточных органелл эукариот — хлоропластов и митохондрий. В составе хлоропластной ДНК обнаружены гены, гомологичные генам, кодирующим а-, - и -субъединицы РНК-полимеразы Е. oli. Это, а также сходство нуклеотидной последовательности промоторов бактерий и хлоропластов свидетельствует о том, что РНК-полимераза хлоропластов должна быть сходна с РНК-полимеразой бактерий. РНК-полимеразы митохондрий состоят, по-видимому, всего из одной субъединицы, подобно РНК-полимеразам, кодируемым некоторыми бактериофагами, такими, как ТЗ и Т7. РНК-полимераза митохондрий дрожжей сходна с РНК-полнмеразами этих фагов по аминокислотной последовательности. Ген, кодирующий митохондриальную РНК-полимеразу, располагается в ядре. [c.136]
Митохотгдриями, или хондриосомами, называются органоиды клетки эукариотов, представляющие собой мембранные внутриклеточные образования. Форма и размеры их различны — от овальных и грущевидных телец до нитевидных или ветвистых. Наиболее полно митохондрии изучены у дрожжей и дрожжеподобных грибов в работах М. Н. Мейселя с сотр. [175, 176], в которых показано, что они не отличаются от митохондрий высших организмов. По своему назначению митохондрии представляют собой центры сосредоточения ферментов энергетического обмена. М. Н. Мейсель обнаружил [175], что клетки дрожжей при брожении содержат меньшее число митохондрий, которые гипертрофированы (бродильный тип клеток). Аналогичная картина наблюдается и в аэробных условиях при избытке углеводов в среде, особенно сахарозы и глюкозы. Цитологически наблюдается бродильная перестройка митохондриального аппарата. [c.71]
Биологический смысл, заключенный в гомологии последовательностей, лучше всего можно проиллюстрировать на примере цитохрома с-железосодержащего митохондриального белка, участвующего в качестве переносчика электронов в процессах биологического окисления в эукариотических клетках. Молекулярная масса этого белка у большинства видов составляет около 12 500 при этом его полипептидная цепь содержит 100 или несколько большее число аминокислотных остатков. Бьии установлены аминокислотные последовательности для цитохромов с, выделенных более чем из 60 видов, и во всех исследованных белках 27 положений в полипептидной цепи оказались занятыми одинаковыми аминокислотными остатками (рис. 6-14). Это указывает на то, что все эти остатки играют важную роль в определении биологической активности цитохрома с. В других положениях аминокислотные остатки могут варьировать от вида к виду. Второй важный вывод, сделанный на основе анализа аминокислотных последовательностей цитохромов с, состоит в том, что число остатков, по которым различаются цитохромы с любых двух видов, пропорционально филогенетическому различию между данными видами. Например, молекулы цитохромов с лошади и дрожжей (эволюционно весьма далеких видов) различаются по 48 аминокислотным остаткам, тогда как цитохромы с гораздо более близких видов— курицы и утки-только по двум остаткам. Что же касается цитохромов с курицы и индейки, то они имеют идентичные аминокислотные последовательности. Идентичны также цитохромы с свиньи, коровы и овцы. Сведения о числе различий в аминокислотных последовательностях гомологичных белков из разных видов используют для построения эволюционных карт, отражающих последовательные этапы возникновения и развития различных видов животных и растений в процессе эволюции (рис. 6-14). [c.155]
Pu . 9.13. Электронная микрофотография митохондриальной ДНК из пивных дрожжей Sa haromy es arlsbergensis. Молекула представляет собой суперспи-рализованное кольцо ДНК с длиной окружности 26 мкм. Она построена примерно из 75 ООО нуклеотидов. В ней закодированы некоторые митохондриальные белки. Прочие необходимые для митохондрий гены находятся в ядерной ДНК. [c.359]
До сих пор мы говорили в основном о бактериальных системах. Если мы обратимся к клеткам животных, дрожжей, Neu-rospora и т. д. (обычно называемым эукариотическими клетками), то обнаружим, что они содержат два типа белоксинтези-рующих систем митохондриальную и цитоплазматическую. Митохондриальная система — это по существу система прокариотического типа, она весьма сходна с системой бактерий действительно, ферменты митохондрий и ферменты бактерий, по-ви-димому, могут полностью заменять друг друга при функционировании рибосом из обоих источников, кроме того, рибосомы митохондрий и прокариот сходны по размеру. С другой стороны, цитоплазматическая белоксинтезирующая система (которую мы называем эукариотической) отличается рядом особенностей, хотя механизм синтеза белка в этой системе и в системе прокариот в общих чертах сходен рибосомы эукариот крупнее [c.59]
Все классы генов могут иметь прерывистое строение все гены, кодирующие белки, а также гены, кодирующие рРНК, и гены, кодирующие тРНК. Интроны обнаружены также в митохондриальных генах дрожжей и хлоропластных генах. Прерывистые гены, по-видимому, присутствуют в клетках эукариот всех классов, хотя их содержание варьирует. Например, их доля среди ядерных структурных генов позвоночных может превышать таковую у грибов. [c.253]
Было обнаружено существование интересного сходства между митохондриальными интронами дрожжей и некоторыми генами рРНК. В их состав входит несколько довольно коротких сходных последовательностей. Они расположены на некотором расстоянии от границ экзон—интрон (на самих границах таких консервативных последовательностей нет). Некоторые последовательности причастны к сплайсингу, по крайней мере, в случае гена box, поскольку они обеспечивают создание сайтов, соответствующих 1/ш -мутациям box 9 и box 2, блокирующим сплайсинг. [c.260]
По-видимому, все митохондриальные интроны дрожжей имеют такие канонические последовательности. Так, их обнаруживают в интронах, имеющих открытые кодирующие участки, и в других интронах, в которых все возможные рамки считывания заблокированы. К ядерным генам, содержащим канонические последовательности, относится ген Tetrahymena, кодирующий предшественник рРНК, подвергающийся автономному сплайсингу. Эти данные служат замечательной демонстрацией существования неожиданного эволюционного сходства ядерного и внеядерных геномов и позволяют сделать предположение о том, что, возможно, сплайсинг разных РНК протекает в соответствии с общим механизмов. Это заключение наводит на мысль об интересном парадоксе, касающемся механизмов сплайсинга. [c.261]
Соответствие по крайней мере некоторых экзонов белковым доменам подтверждает предположение о том, что оно имеет фундаментальное значение в эволюции генов. Ясно, что дупликации и слияние экзонов могли играть важную роль в эволюции. Мы не можем проследить за действительными событиями, проишедшими в ходе эволюции каждого гена. Имеется несколько примеров взаимоотношений между экзонами и белковыми доменами, когда отсутствует их простое соответствие, но это можно объяснить тем, что такие события, как слияние экзонов, изменили структуру гена-предка в процессе эволюции ядерных генов. Однако в ряде случаев мы сталкиваемся с большими несоответствиями между структурами генов и белков. Митохондриальные гены дрожжей и млекопитающих кодируют практически идентичные митохондриальные белки, несмотря на существенные различия в организации генов. Геном митохондрий позвоночных очень мал и имеет чрезвычайно компактную организацию нерасщенленных генов (гл. 22), тогда как митохондриальный геном дрожжей имеет большие размеры и включает ряд сложных прерывистых генов. Какая форма гена была исходной [c.265]
Геном митохондрий дрожжей существенно больше. Его размер у различных штаммов S. erevisiae варьирует в широких пределах, составляя в среднем около 84 т. п. н. В дрожжевых клетках при стандартном определении обнаруживается около 22 митохондрий на одну клетку, что предполагает наличие примерно 4 копий генома в каждой органелле. В растущих клетках содержание митохондриальной ДНК может достигать 18% всей ДНК клетки. (Митохондриальные геномы других видов дрожжей могут иметь существенно большие размеры.) [c.282]
Данные о происхождении компонентов, необходимых для экспрессии генов в митохондриях дрожжей, приведены в табл. 22.3. В целом аналогичная картина наблюдается в митохондриях других видов организмов и в хлоропластах. В митохондриях дрожжевых клеток можно выделить восемь мРНК, и они транскрибируются с помощью РНК-полимеразы, которая (предположительно) кодируется ядерными генами. Только мРНК, транскрибируемые с митохондриальных генов, могут транслироваться внутри митохондрии, и, с другой стороны, это единственное место в клетке, где эти мРНК могут экспрессироваться. [c.283]
В табл. 22.4 приведены комплексы, содержащие белки, синтезированные в митохондриях дрожжей. АТРаза состоит из двух частей мембранного фактора, образуемого двумя или более субъединицами, кодируемых митохондриальным геномом, и растворимой АТРазы F1, состоящей примерно из пяти субъединиц, синтезируемых в цитоплазме. Цитохром-с—оксидаза также состоит из субъединиц, происходящих из обоих источников. В состав комплекса цитохромов Ьс входит один белок митохондриального происхождения, связанный с щестью субъеди- ницами цитоплазматического происхождения. Малая субъединица рибосомы включает в себя один белок (Уаг 1), кодируемый митохондриальными генами. Были получены мутации, позволяющие идентифицировать почти все митохондриальные гены. [c.284]
Существование генетической рекомбинации в митохондриях дрожжей (см. ниже) позволило создать генетическую карту генома митохондрий, которая была сопоставлена с физической картой при изучении возникающих естественным путем делеций митохондриального генома (называемых петит-мутациями). Участок, занимаемый каждым геном, определяли по положению на карте соответствующей мРНК. [c.285]
На рис. 22.2 приведена карта митохондриального генома дрожжей, показывающая взаимное расположение генов для основной части РНК и белков (за исключением молекул тРНК, число видов которьа составляет около 22 и гены для которых до сих пор полностью не картированы). Наиболее характерная особенность этой карты — пространственная разделенность локусов. [c.285]
Отличительная особенность митохондриального генома дрожжей состоит в пространственной разобщенности генов, кодирующих рРНК, что встречается крайне редко. Ген, кодирующий 15S-pPHK, непрерывен и располагается на расстоянии около 25 000 п.н. от гена для 21S-pPHK. В некоторых штаммах дрожжей последний ген имеет один интрон (как показано на карте), в других штаммах он не прерывается. [c.285]
Весьма значительная часть митохондриального генома дрожжей (около 25%) состоит из коротких (А—Т)-бо-гатых участков ДНК, вероятно не несущих функции кодирования. Однако значительная часть генетического материала все еще не изучена, и будет удивительно, если для оставшихся незаполненными участков карты не будут обнаружены другие гены. Но, даже допуская такую возможность, можно все же считать, что общее число генов в митохондриях дрожжей вряд ли превышает 20. [c.285]
У дрожжей S. erevisiae рекомбинация между митохондриальными маркерными генами-обычное явление. Природа такой рекомбинации не известна, но кроссинговер между двумя родительскими митохондриальными ДНК, имеющими значительные различия, приводит к появлению потомства, обладающего рекомбинантными ДНК. Этот факт непосредственно свидетельствует о том, что митохондриальные ДНК обеих родительских особей вступают в контакт. [c.287]
Все петит-мутации нарушают функционирование митохондрий. Они не становятся летальными, поскольку дрожжи могут существовать и в аэробных условиях (когда необходимо дыхание) и в анаэробных (когда без него можно обойтись). Следовательно, мутации митохондриального генома являются условно-летальными, вызывающими гибель дрожжей лишь в аэробных условиях в анаэробных же условиях они выживают. Таким образом, нарушение функций митохондрий приводит к переходу на анаэробный способ существования. Несомненно, такой переход невозможен в случае, например, клеток животных, для которых прекращение функционирования митохондрий оказывается летальным. (Аналогичная генетическая ситуация наблюдается в случае хлоропластов С. reinhardii, которые не нуждаются в фотосинтезе в присутствии ацетата.) [c.288]
chem21.info





















 |  |  |  |  |  |  |  |  |  |  |  |  |  |  |
 Пример видео 3 Пример видео 3 |  Пример видео 2 Пример видео 2 |  Пример видео 6 Пример видео 6 |  Пример видео 1 Пример видео 1 |  Пример видео 5 Пример видео 5 |  Пример видео 4 Пример видео 4 |
Администрация муниципального образования «Городское поселение – г.Осташков»