



Справочник химика 21. В клетках дрожжей не обнаружено
Модификации в клетках дрожжей - Справочник химика 21
Эффективность экспрессии белков человека в дрожжевых системах зависит от природы самого продукта известны успешные опыты, есть здесь и не очень рекламируемые неудачи (как, например, i-интерферон, на долю которого приходится менее 0,1% всех белков, образующихся в дрожжевой клетке). Вдобавок существуют белки — например, многие из тех, что участвуют в процессах свертывания крови, — посттрансляцион-ная модификация которых происходит по такому пути, который не может реализоваться в клетках дрожжей (в данном случае это гамма-карбоксилирование остатков глутаминовой кислоты) [7]. [c.210]
Многие биологически активные белки высших эукариот синтезируются в форме предшественников, подвергающихся секреции из клеток. В ряде экспериментов бьшо обнаружено, что если белки такого типа (например, гормон роста быка, активатор тканевого плазминогена человека, у-интерферон человека, прохимозин быка) в процессе синтеза в бактериальной или дрожжевой клетке остаются в цитоплазме, то они переходят в нерастворимую и неактивную форму. Поэтому создание продуцентов, обеспечивающих секрецию и сопутствующую ей модификацию белка, является крайне важной задачей генетической инженерии и биотехнологии. Большое внимание уделяется дрожжам, у которых ряд белков эффективно выводится из клеток в окружающую среду. В отличие от бактерий в клетках дрожжей в процессе секреции может происходить гликозилирование и правильная укладка эукариотических белков. Особо следует отметить, что большинство штаммов S. erevisiae не вьщеляет в культуральную среду протеазы, что в еще большей степени повышает перспективность данной системы для создания высокопродуктивных штаммов, секретирующих в окружающую среду целевые белки. [c.318]
Нередко возникает задача ввести ген в клетки эукариот, например в дрожжевые клетки, в которых могут нарабатываться белки, прошедшие после их образования необходимые стадии модификации, несвойственные прокариотическим клеткам. Для этой цели используют специальные, так называемые челночные, векторы, которые могут автономтю размножаться как в прокариотических, так и в эукариотических клетках, например в Е.соН и дрожжах. В эукариотические клетки плазмиды вводят на заключительных стадиях, поскапьку многие предварительные этапы клонирования существенно проще проводить в кле гках прокариот. [c.304]
Помимо химических средств, гипотермии и гипоксии модификация радиорезистентности производилась с помощью фракционированного облучения в невысоких летальных дозах. На клетках асцитной карциномы было показано, что при увеличении времени между двукратным облучением одновременно с возрастанием радиорезистентности объектов общее содержание эндогенных тиолов увеличивается. Параллельное исследование радиорезистентности и уровня сульфгидрильных групп проводилось также на клетках, находящихся на разных стадиях роста и клеточного цикла. Так, Э. Я. Граевский (1969) привел сравнительные данные из работ по изучению изменения тиолов и радиорезистентности микроспор в процессе клеточного деления. Оказалось, что в процессе мейоза и митоза происходят однонаправленные изменения содержания тиолов в клетках и их устойчивости (устойчивость оценивалась по выходу хромосомных аберраций) к действию ионизирующей радиации. Динамика изменения уровня эндогенных сульфгидрильных групп в зависимости от изменения радиорезистентности прослежена также на синхронно делящейся икре морских ежей в различных стадиях клеточного цикла, на растущих клетках асцитной карциномы Эрлиха в процессе ее старения, на синхронной культуре клеток разных штаммов хлореллы в процессе клеточного деления, на клетках в различных фазах роста. Эти данные позволили авторам заключить, что изменения радиочувствительности в цикле связаны не только с изменением генетического аппарата в клетке, но и с варьированием содержания внутриклеточных тиолов, выполняющих функции эндогенных радиопротекторов. Эти представления получили дополнительное обоснование в работе Ю. В. Корогодиной и др. (1975). Так, на диплоидных дрожжах (штамм Мегри 139 В) было установлено, что клетки, находящиеся в логарифмической фазе роста, в отличие от стационарной фазы более радиорезистентны и содержат в полтора раза больше сульфгидрильных групп. Авторы считают, что именно высокий уровень тиолов почкующихся дрожжевых клеток может определять их повышенную радиорезистентность. [c.283]
Конверсия генов. Еще один относящийся к обсуждаемому предмету феномен давно известен в экспериментальной генетике под названием генной конверсии [122]. Различные данные, полученные при изучении глобиновых генов, позволяют предполагать наличие такого феномена и в геноме человека (разд. 4.3 см. также рис. 2.97). Генная конверсия есть не что иное, как модификация одного из двух аллелей другим, в результате чего гетерозигота Аа, например, становится гомозиготой АА. Винклер, который впервые обсуждал этот феномен более 50 лет тому назад, допускал физиологическое взаимодействие аллелей. Однако работы на дрожжах показали, что он связан с атипичной рекомбинацией. Данный процесс иллюстрирован на рис. 2.97. Кроссинговер всегда приводит к разрыву последовательности ДНК в сайте перекреста. Обычно разрыв репарируется, для чего последовательность сестринской хроматиды используется как матрица. Таким образом восстанавливается исходная двойная спираль. Однако иногда репарация осуществляется на матрице гомологичной хромосомы. В этом случае наблюдаются отклонения от обычной сегрегации. Генная конверсия имеет место и в соматических тканях, особенно у растений. Возможно, что в этом случае рекомбинационный процесс протекает атипично. Наличие генной конверсии не является неожиданным, поскольку спаривание гомологичных хромосом в соматических клетках и соматический кроссинговер характерны для многих видов [c.144]
На устойчивость микроорганизмов к процессам криоконсервирования влияют самые различные факторы возраст клеток, условия их культивирования, плотность клеток в суспензии. Установлено, в частности, что клетки Е. соИ проявляют наибольшую криорезистентность в конце логарифмической или в начале стационарной фазы роста. Культивирование дрожжей и молочнокислых бактерий в средах, содержащих ненасыщенные жирные кислоты, повышает их устойчивость к замораживанию. Эти изменения криорезистентности клеток обычно связывают с модификацией мембранных структур. Что касается плотности клеток, то установленный факт повышения титра жизнеспособных микроорганизмов с увеличением исходной концентрации клеток в суспензии пока не имеет объяснения. [c.73]
У представителя низших эукариот Sa kammy es erevisiae имеется по крайней мере пять генов, кодирующих белки, которые участвуют во внесении разрывов в УФ-облученную ДНК. Нарушение только в одном из этих пяти ЛЛЛ-генов приводит к тому, что клетки утрачивают способность к внесению разрывов в ДНК и, следовательно, к удалению пиримидиновых димеров. У дрожжей существуют также мутанты с нарушенной способностью к удалению сшивок между цепями, хотя элиминация УФ-индуцированных повреждений проходит нормально. Это предполагает, что у дрожжей, как и у человека, для удаления поперечных сшивок, а возможно, и для исправления множества других химических модификаций в ДНК существуют специфические, весьма сложные механизмы репарации. [c.102]
chem21.info
Брожение в дрожжевых клетках, формы
К аскомицетам относятся также дрожжи, или дрожжевые грибы. Это одноклеточные организмы, размножающиеся вегетативно — почкованием (например, сахаромицеты) и делением (шизосахаромицеты) или половым путем с образованием внутри клетки аскоспор. Форма клетки круглая, овальная или палочковидная (рис. 26). Клетки крупные — до 10 мкм. При выращивании в жидкой среде дрожжи могут образовывать пленку на поверхности или расти в глубине среды, вызывая в последнем случае брожение углеводов. На твердой среде дрожжи образуют колонии характерной для каждого вида формы, структуры, цвета и консистенции. Кроме истинных дрожжей, или аскомицетов, есть также ложные или аспорогенные дрожжи, не образующие спор. Дрожжи и дрожжеподобные грибы широко распространены в природе и встречаются в почве, на поверхности растений и насекомых. [c.76] Молодые клетки дрожжей, размноженных на пивном сусле, имеют прозрачную плазму, хороший тургор, образуют цепочки из нескольких клеток. При размножении в подкисленном сусле в плазме клеток можно обнаружить зернистость, клетки более вытянуты, цепочки не образуются, дрожжи распылены. К концу брожения в зрелых дрожжах плазма клеток уплотнена, изредка встречаются. мертвые клетки. Слишком высокая кислотность среды приводит к тому, что клетки мельчают, принимают неправильную форму. Дрожжи, переведенные в бродильный чан, уже через несколько часов брожения меняют вид, становятся крупнее, почкуются, плазма делается гомогенной, вакуоль разбивается на несколько пузырьков. Во время дображивания и в зрелой бражке плазма клеток уплотняется, появляются большие вакуоли, часть клеток отмирает. При известном навыке морфологический вид дрожжей дает возможность сделать заключение о их физиологическом состоянии. [c.233]В анаэробных условиях грибы растут, как правило, очень плохо и лишь короткое время. В отсутствие молекулярного кислорода они переходят к брожению многие из них образуют в таких случаях молочную кислоту или этиловый спирт. Изменяется при этом и форма роста Мисог ra emosus, например, образует в анаэробных условиях почкующийся мицелий, и его молодые клетки, подобно дрожжам, размножаются почкованием. [c.62]
Митохотгдриями, или хондриосомами, называются органоиды клетки эукариотов, представляющие собой мембранные внутриклеточные образования. Форма и размеры их различны — от овальных и грущевидных телец до нитевидных или ветвистых. Наиболее полно митохондрии изучены у дрожжей и дрожжеподобных грибов в работах М. Н. Мейселя с сотр. [175, 176], в которых показано, что они не отличаются от митохондрий высших организмов. По своему назначению митохондрии представляют собой центры сосредоточения ферментов энергетического обмена. М. Н. Мейсель обнаружил [175], что клетки дрожжей при брожении содержат меньшее число митохондрий, которые гипертрофированы (бродильный тип клеток). Аналогичная картина наблюдается и в аэробных условиях при избытке углеводов в среде, особенно сахарозы и глюкозы. Цитологически наблюдается бродильная перестройка митохондриального аппарата. [c.71]
Препарат обладает побочным действием против оидиума винограда, различных видов пенициллиума, мучнистой росы и парши яблони. Подавляет возбудителей болезней, особенно В. inerea, которые приобрели резистентность к производным бензимидазола, а также устойчивых к ртутьсодержащим препаратам форм возбудителя гельминтоспориоза. При обработке винограда в борьбе с серой гнилью ровраль не оказывает воздействия на спиртовое брожение, хотя препарат поглощается клетками дрожжей, участвующих в ферментащ1И вина. [c.113]
Основной органической кислотой, выделяемой в питательную среду, является уксусная. Она образуется в процессе окисления ацетальдегида путем удаления атомов водорода в ходе реакции, обратной обычному восстановлению ацетальдегида до этилового спирта. Скорость образования уксусной кислоты можно повысить путем добавления в питательную среду шелочей, стимулирующих выработку дрожжами уксусной кислоты и регулирующих значение pH среды. В ходе большинства процессов дрожжевого брожения pH среды снижается до значений 3,5-4. Среднецепочечные жирные кислоты, попадая в питательную среду, придают конечному продукту неприятный козий привкус, однако жирные кислоты с более высокой молекулярной массой обычно включаются в структуру клетки в форме фосфолипидов. По имеющимся данным, продуцирование жирных кислот стимулируется повышенным содержанием в питательной среде фосфора, магния и биотина [21]. [c.57]
Биохимический метод получения оптически активных соединений наряду с методом механического отбора (разд. 4-4а) и методом разделения, основанным на кристаллизации диастереомеров (разд. 4-46), был впервые применен Пастером [68]. Пастер заметил, что при брожении аммонийной соли рацемической винной кислоты, вызванном дрожжами или Peni illium glau um (грибковой плесенью), расходовалась преимущественно природная (правовращающая) форма и по истечении некоторого времени из подвергавшейся ферментации жидкости можно выделить соль чистой (—)-винной кислоты. Очевидно, это случай асимметрической деструкции грибковая плесень потребляет для своей жизнедеятельности преимущественно природный энантиомер и оставляет нетронутым другой, не встречающийся в природе (левовращающий) энантиомер. В более поздней работе, посвященной синтезу нитрила миндальной кислоты в присутствии эмульсина (см. выше), было ясно показано, что присутствие живого организма или живой клетки не обязательно для асимметрического синтеза этого типа и что активными агентами являются ферменты (которые могут функционировать как внутри, так и вне живого организма). [c.78]
chem21.info
Дикие дрожжи (часть 2). Микробиология и биохимия вина.
Дикие дрожжи (часть 2)
Hansenula anomala (ганзенула аномала) – клетки этих дрожжей различны по величине и форме (рис.10). Их размеры (1,5-6,0) X (3,0-30) мкм. Клетки, образующие пленку, удлиненные, с большими вакуолями и жировыми включениями, сильно преломляющими свет. Клетки осадка – меньшего размера, овальной или округлой формы с однородной плазмой. Располагаются в виде одиночных клеток, парами или цепочками. Образуемые ими споры имеют характерную шляповидную форму и содержатся от 1 до 4 в аске.

Рис.10. Дрожжи Hansenula (Х2000).
Дрожжи Наnsеnulа аnоmala сбраживают глюкозу, маннозу, сахарозу, галактозу, мальтозу. Они быстро развиваются на поверхности виноградного и плодового сусла, образуя на вторые сутки пленку, на третьи - осадок, вызывая одновременно брожение с образованием 2-3% об. спирта. Большинство штаммов может развиваться на поверхности вина с содержанием спирта не выше 10% об. и только некоторые - при 11-13% об., образуя при этом сухую матовую пленку серовато-белого цвета, высоко ползущую по стенкам сосуда.
Hansenula является энергичным эфирообразователем. Она сообщает вину сильный, но несвойственный ему аромат, обусловленный образованием этил ацетата. Летучие кислоты образует в небольшом количестве. Hansenula вместе с Pichia и Candida является возбудителем болезни - цвели вина, может также вызвать дрожжевые помутнения столовых вин.
Candida mycoderma (кандида микодерма) - клетки этих дрожжей имеют овальную или цилиндрическую форму с закругленными концами (рис.11). Размер клеток (2-4) Х (3,5-9,0) мкм. Жировые отложения накапливаются в клетках в виде шариков и нередко биполярно (в противоположных концах клетки), сильно преломляют свет. В сусле и вине имеет вид сухой матовой морщинистой пленки серовато-белого цвета.
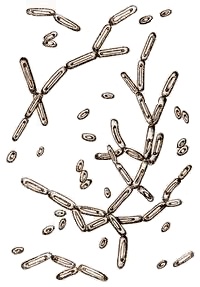
Рис.11. Дрожжи Candida mycoderma (Х800).
Спор С. mycoderma не образует, брожения не вызывает. Развиваясь на поверхности вина при доступе воздуха, С. mусоderma снижает содержание в вине спирта и экстракта, обогащая его летучими кислотами, придающими вину острый вкус. При глубоко зашедшем процессе спирт может полностью разложиться.
С. mycoderma является главным возбудителем болезни вина - цвели. Продукты ее обмена в кислой среде (летучие кислоты и др.) задерживают развитие винных дрожжей и снижают их бродильную способность при вторичном брожении.
Torulopsis (торулопсис) - к этому роду относятся почкующиеся одноклеточные организмы, у которых спорообразование не обнаружено. Форма клеток круглая, реже овальная. Величина клеток (2,9-6,5) Х (2,9-7,2) мкм. В клетках накапливается большое количество жира, сильно преломляющего свет, в виде капелек, заполняющих всю клетку. В сусле и вине появляется слизь.
Характерная особенность этих дрожжей заключается в одновременном образовании нескольких почек в различных частях материнской клетки. В зависимости от вида Torulopsis обладают различной бродильной способностью - от слабой до сильной.
Вид Torulopsis bacillaris (бациллярис) вызывает самопроизвольное брожение сусел из белых сортов винограда, при этом накапливается 7-10% об. спирта.
Brettanomyces (бреттаномицес) - эти дрожжи широко распространены в природе. При брожении они образуют эфиры, обладающие фруктовым (яблочным) ароматом, поэтому они используются для дополнительного брожения определенных сортов английского (британского) пива.
Клетки по форме разнообразные - круглые, овальные, стрельчатые, часто заостренные с одной или двух сторон (рис. 12). Встречаются и колбасовидные клетки. Размеры (2,5-8,8) Х (3,1-6,8) мкм. По сравнению с винными дрожжами они бродят медленно и образуют 9-12% об. спирта. При закрытом брожении они могут образовать 13% об. спирта, но 14 % об. уже действуют на них губительно.
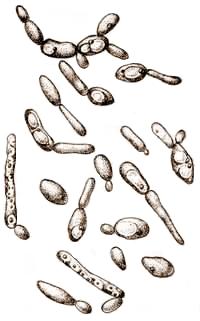
Рис 12. Дрожжи Вrettanomyсes (Х1500).
Некоторые штаммы Brettanomyces способны при определенных условиях образовывать пленку, а затем вызывать брожение. Другие же, наоборот, сначала сбраживают сахар, а уже после этого образуют пленку. При развитии в виноградном сусле и вине дрожжи Brettanomyces могут вызвать их порчу. При этом сусло и вино мутнеют, приобретают несвойственный им яблочный тон, в них накапливается значительное количество уксусной кислоты. Особенно большой вред эти дрожжи могут принести шампанскому производству, вызывая недоброд тиражной смеси, образуя пылевидный осадок, что затрудняет сведение его на пробку и отделение, повышая содержание летучих кислот, придающих шампанскому фруктовые тона и значительно ухудшающих игристые свойства.
часть 1 <<< часть 2
vinobio.narod.ru





















 |  |  |  |  |  |  |  |  |  |  |  |  |  |  |
 Пример видео 3 Пример видео 3 |  Пример видео 2 Пример видео 2 |  Пример видео 6 Пример видео 6 |  Пример видео 1 Пример видео 1 |  Пример видео 5 Пример видео 5 |  Пример видео 4 Пример видео 4 |
Администрация муниципального образования «Городское поселение – г.Осташков»