



Большая Энциклопедия Нефти и Газа. Клетки дрожжи
4. Дрожжевая клетка. Цитология. Биология дрожжей
Похожие главы из других работ:
Античный период в истории естествознания. Состав и строение клетки
2. Клетка как структурная и функциональная единица живого. Состав и строение клетки
Современная клеточная теория включает следующие положения: 1. Все живые организмы состоят из клеток. Клетка - структурная, функциональная единица живого, основная единица строения и развития всех живых организмов...
Ионные механизмы потенциала покоя
Идеальная клетка
Начнем с рассмотрения модели идеальной клетки, показанной на рис.1.1. В этой клетке содержатся ионы калия, натрия и хлора, а также крупные анионы. Клетка помещена в раствор, содержащий ионы натрия и калия. Другие ионы...
Общая биология
Клетка, как открытая система. Организация потоков веществ и энергии. Биологическое окисление, дыхание, брожение. Фото- и хемосинтез
Обязательным условием существования любого организма является постоянный приток питательных веществ и постоянное выделение конечных продуктов химических реакций, происходящих в клетках...
Объекты биотехнологии в пищевой промышленности
Глава 2. Клетка как основной объект биотехнологических исследований
1) Живым организмам свойственные общие принципы структуры: . - единство элементарного состава; . - единство типов химических соединений; . - единство субклеточной организации; V. - единство клеточного строения...
Основы биологии
Лекция 2. Клетка - элементарная структурная единица живого организма
План 1. Клеточная теория 2. Строение клетки 3. Эволюция клетки Клеточная теория В 1665г. Р. Гук впервые обнаружил растительные клетки. В 1674 г. А. Левенгук открыл животную клетку. В 1839 г. Т. Шванн и М .Шлейден сформулировали клеточную теорию...
Основы естествознания
1. Клетка - структурная и функциональная единица живой материи
Клеточная теория в современном виде включает три главных положения. Первое положение соотносит клетку с живой природой планеты в целом. Оно утверждает, что жизнь, какие бы сложные или простые (например, вирусы) формы она ни принимала...
Особенности строения клетки
1. Клетка как элементарная структурная единица организма. Основные компоненты клетки
Клетка - основная структурная и функциональная единица жизни, ограниченная полупроницаемой мембраной и способная к самовоспроизведению. В растительной клетке, прежде всего, нужно различать клеточную оболочку и содержимое...
Продукты питания и процессы старения организма человека
4. Старение тканей. Как старится клетка
Изучение старения тканей связано с именем академика А.Н. Богомольца, рассматривавшего соединительную ткань как своего рода мягкий скелет организма, который с возрастом уплотняется и "окутывает" клетки других тканей...
Скелет человека
4 Грудная клетка
Грудная клетка состоит из ребер, соединенных передними концами с грудиной, а задними -- с грудными позвонками. Фронтальная поверхность грудной клетки, представленная грудиной и передними концами ребер, значительно более короткая...
Строение грудной клетки
1.4 Грудная клетка
Грудную клетку, compares thoracis, составляют грудной отдел позвоночного столба, ребра (12 пар) и грудина. Грудная клетка образует грудную полость, cavitas thoracis, имеющую форму усеченного конуса, обращенного широким основанием книзу...
Строение и функции опорно-двигательного аппарата человека
2.3 Грудная клетка
Грудная клетка представляет собой костно-хрящевое образование, состоящее из грудных позвонков, 12 пар ребер и грудины, соединенных между собой при помощи различных видов соединений. У грудной клетки различают 4 стенки (переднюю...
Структурные уровни живого
2. Клетка как «первокирпичик» живого.
Фундаментальная частица в биологии - живая клетка. Именно она является мельчайшей системой, обладающей всем комплексом свойств живого, в том числе и носителем генетической информации...
bio.bobrodobro.ru
Обмен веществ дрожжевой клетки
⇐ ПредыдущаяСтр 16 из 169Следующая ⇒Для осуществления жизненно необходимых процессов обмена веществ и для образования новой клеточной субстанции дрожжи, подобно любым другим клеткам, нуждаются в энергии и питательных веществах. Энергию для поддержания этих процессов дрожжи, как и все другие живые существа, добывают главным образом с помощью дыхания. Получение энергии при дыхании весьма значительно, поскольку глюкоза разлагается без остатка. При дыхании полученные питательные вещества (например, сахара) полностью расщепляются на СО2 и воду:
С6Н12О6 + 6О2 → 6Н2О + 6СО2.
При отсутствии воздуха дрожжи (единственный живой организм) переходят на спиртовое брожение. При этом из глюкозы образуется спирт (этанол) и СО2:
С6Н12О6 → 12C5H5OH + 2СО2.
Путем брожения дрожжевая клетка получает несравнимо меньше энергии, чем путем дыхания (см. также раздел 4.1).
Расщепление глюкозы до спирта или - при дыхании - до СО2 и воды проводится путем большого числа последовательных реакции. При этом каждая из них осуществляется с помощью специального фермента-катализатора. В дрожжевой клетке эти ферменты связаны с определенными клеточными структурами. Так, ферменты для глюкозы и спиртового брожения находятся в цитоплазме, тогда как дыхание осуществляется благодаря ферментам, локализованным в митохондриях.
Органические вещества, необходимые для процесса дыхания и для сбраживания, поглощаются интегрированными протеинами клеточной мембраны и транспортируются через нее. Поэтому дрожжевой клеткой могут быть поглощены лишь такие вещества, для которых имеются соответствующие транспортные механизмы. Это, в свою очередь, зависит от количества и типов ферментов дрожжевой клетки.
Клетка осуществляет комплексный обмен веществ, а именно:
· углеводный;
· белковый;
· жировой;
· обмен минеральных веществ.
Углеводный обмен веществ служит прежде всего для получения энергии путем дыхания и брожения, при этом лишь небольшая часть содержащихся в сусле сахаров откладывается в качестве резерва в виде гликогена и трегалозы.
Белковый обмен веществ так же. как обмен жировых и минеральных веществ, служит, в первую очередь, для построения новых клеточных субстанций, причем здесь существенную роль играют как процессы их построения, так и процессы распада. Эти очень сложные процессы обмена веществ, оказывающие решающее влияние на качество пива, будут более подробно рассмотрены в разделе 4.1.2.
Размножение и рост дрожжей
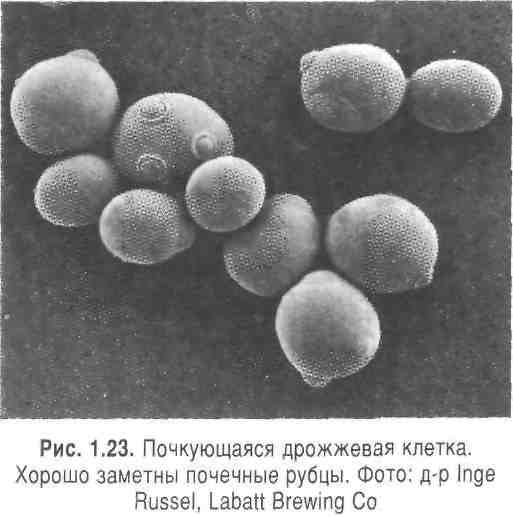 Типичным способом размножения дрожжей является почкование, и поэтому их еще называют почкующимися грибами. При почковании из материнской клетки образуется маленькая пузырьковая выпуклость, в которую переходит часть цитоплазмы, а также дочернее ядро, образующееся путем деления. У одних некоторых штаммов дрожжей материнская и дочерняя клетки отделяются друг от друга, причем на материнской клетке остается почечный рубец (рис. 1.23). У других штаммов клетки остаются взаимосвязанными и образуют почечные сообщества.
Типичным способом размножения дрожжей является почкование, и поэтому их еще называют почкующимися грибами. При почковании из материнской клетки образуется маленькая пузырьковая выпуклость, в которую переходит часть цитоплазмы, а также дочернее ядро, образующееся путем деления. У одних некоторых штаммов дрожжей материнская и дочерняя клетки отделяются друг от друга, причем на материнской клетке остается почечный рубец (рис. 1.23). У других штаммов клетки остаются взаимосвязанными и образуют почечные сообщества.

Если микроорганизмы перенести в свежий питательный раствор, как это происходит, например, при засеве сусла дрожжами на пивоваренном заводе, то последние начинают расти. Этот рост характеризуется определенными закономерностями - различают шесть фаз роста, протекающих с различной скоростью (рис. 1.24).
Латентная, или индукционная фаза.В этой фазе, называемой также фазой разбега, происходит активизация обмена веществ. Длительность этой фазы сильно варьирует. Она зависит от вида микроорганизма, возраста культуры и от условий выращивания. Эта фаза, называемая также лаг-фаза (lag-phase), заканчивается с началом деления клеток. («lag» по-английски - отставание, задерживание - Прим. ред.)
Фаза ускорения.В этой фазе, примыкающей к латентной, скорость деления клеток возрастает с ускорением.
Экспоненциальная фаза.В этой фазе экспоненциального или логарифмического размножения, для краткости называемой лог-фазой (log-phase), скорость размножения постоянна и максимальна. Время генерации, то есть отрезок времени, за которое число клеток удваивается, достигает в этой фазе своего минимума. Для дрожжей в типичных для пивоварения условиях размножения он составляет несколько часов.
Фаза замедления. В результате действия различных факторов - например, обеднения субстрата питательными веществами или насыщения тормозящими рост продуктами обмена веществ - лог-фаза ограничена по времени и переходит в фазу замедления с убывающей скоростью размножения.
Стационарная фаза.В заключительной, стационарной фазе число микроорганизмов остается постоянным. Устанавливается равновесие между числом вновь образующихся и отмирающих клеток.
Фаза отмирания.В этой, последней, фазе число погибающих клеток превышает число образующихся путем размножения, и общее число клеток сокращается.
На длительность и интенсивность отдельных фаз роста существенно влияют субстрат, температура и физиологическое состояние дрожжей. Субстрат должен содержать все необходимые для их роста питательные вещества. Так же существенны состав воды, pH и концентрация кислорода в субстрате.
Вода - главная составная часть живой материи и играет в жизнедеятельности микроорганизмов чрезвычайно важную роль. В общем, микроорганизмы могут развиваться только в субстратах, содержащих как минимум 15% воды. Микроорганизмы отличаются друг от друга в отношении оптимального значения pH. Дрожжи предпочитают расти в кислых растворах. О значении снабжения кислородом для роста дрожжей уже было сказано выше. На пивоваренных предприятиях росту дрожжей способствует аэрация сусла перед введением дрожжей.
На рост микроорганизмов решающим образом влияет и температура. Каждый вид микроорганизмов отличается своей оптимальной для развития температурой, при которой латентная фаза и время генерации минимальны. Однако рост дрожжей возможен не только при оптимальной температуре, но может происходить и в более или менее относительно широком диапазоне температур. Для дрожжей рода Saccharomyces она лежит в области температур от 0 до 40°С, а оптимум составляет около 25-30°С.
Физиологическое состояние клеток микроорганизмов, возраст, состояние питательной среды питания - все эти факторы сильно влияют на продолжительность латентной фазы. Очень быстрая активация обмена веществ происходит у тех дрожжевых клеток, которые в экспоненциальной фазе роста были перенесены в свежий субстрат. В условиях пивоваренного предприятия это означает, что быстрое наступление брожения легче всего достичь с теми дрожжами, которые отбираются в стадии главного брожения и без промежуточного хранения вводятся в готовое сусло.
Читайте также:
lektsia.com
Дрожжевая клетка
Дожжевая клетка имеет все основные структуры, которые присущи любой эукариотической клетке, но в то же время она обладает особенностями, свойственными грибам, а именно, сочетанием признаков как растительной, так и животной клеток: клеточная стенка у них ригидная, как у растений, но в клетке отсутствуют хлоропласты и накапливается гликоген, как у животных.
Компоненты дрожжевой клетки
Ядро
В дрожжевой клетке в фазе между делениями всегда имеется только одно ядро. В световом микроскопе его можно увидеть после специальной окраски или с помощью фазово-контрастного устройства при высоких разрешениях. На электронно-микроскопических снимках ультратонких срезов дрожжевых клеток ядро выглядит как более или менее округлая органелла, окруженная двойной мембраной. В ней есть поры в виде округлых сквозных отверстий, которые образуются в результате слияния двух ядерных мембран. Однако ядерные поры - не просто отверстия, они заполнены сложноорганизованными структурами, которые называют комплексом пор ядра. Считается, что основная функция ядерных пор - транспорт готовых рибосомных субъединиц в цитоплазму. Ядерная оболочка многофункциональна, но в основном играет роль барьера, отделяющего содержимое ядра и регулирующего транспорт макромолекул между ядром и цитоплазмой. Основные функциональные единицы ядра - молекулы ДНК, несущие основную генетическую информацию о клетке. ДНК составляет основную часть хроматина - основного компонента ядра. Число хромосом в ядре разных видов дрожжей может быть различным, оно колеблется от 2 до 16.
Митохондрии
В митохондриях имеется собственная митохондриальная ДНК (мДНК), а также весь аппарат белкового синтеза, включая матричную РНК и 70S рибосомы (в отличие от 80S рибосом в цитоплазме). мДНК у дрожжей составляет 5-20% от всей ДНК клетки. Число митохондрий в одной дрожжевой клетке варьирует в пределах 1-20 в разные периоды роста и в зависимости от условий. Как правило, 1-2 митохондрии в клетке более крупные, чем остальные и имеют разветвленную форму. Реконструкция ультратонких срезов клетки позволяет предположить, что в некоторых случаях (в подготовительный период почкования) клетка содержит всего одну вытянутую и сильно разветвленную митохондрию. Митохондрии способны к самовоспроизведению.
Цитоплазаматическая мембрана
На поперечном срезе под электронным микроскопом мембрана у дрожжей выглядит как трехслойная структура. Она представляет собой два слоя фосфолипидов, в которые погружены белковые молекулы, то есть построена по общему для всех клеточных мембран принципу. Однако, имеются различия, касающиеся химического состава. У Saccharomyces cerevisiae основными фосфолипидами мембран являются лецитин, фосфатидилэтаноламин и фосфатидилсерин. На их долю приходится около 90% всех липидов мембраны. В состав мембраны дрожжей входят стероиды - эргостерол, зимостерол и др. Белки представлены в основном ферментами, которые участвуют в трансмембранном переносе веществ, расщеплении полисахаридов и синтезе внеклеточных структур. . Функции цитоплазматической мембраны многообразны: регуляция биосинтеза клеточной стенки, активный транспорт транспорт в клетку специфических молекул органических веществ, транспорт ионов K+ и Na+ и др.
Вакуоли
В фазово-контрастном микроскопе в клетках дрожжей хорошо видны светлые и прозрачные структуры круглой формы. Это вакуоли. Обычно их 1-3 в клетке. Каждая вакуоль окружена одинарной мембраной и содержит различные ферменты, липиды, низкомолекулярные продукты метаболизма (аминокислоты), ионы металлов. В вакуолях сосредоточена большая часть ионов калия.
 Иногда в вакуоли видны «пляшущие» за счет броуновского движения плотные гранулы. Это так называемые метахроматические гранулы, «пляшущие тельца» (dancing bodies), или волютин. Гранулы эти состоят из полимеризованных остатков фосфатов, а по периферии они покрыты комплексными соединениями из РНК, белков и липидов.
Иногда в вакуоли видны «пляшущие» за счет броуновского движения плотные гранулы. Это так называемые метахроматические гранулы, «пляшущие тельца» (dancing bodies), или волютин. Гранулы эти состоят из полимеризованных остатков фосфатов, а по периферии они покрыты комплексными соединениями из РНК, белков и липидов. Волютин - это резерв полифосфатов в клетке. Основная функция вакуолей - разобщение процессов синтеза и распада белков и нуклеиновых кислот. Они выполняют также роль депо для хранения некоторых запасных веществ и ферментов, участвуют в регуляции тургорного давления. Также в клетке присутствуют: клеточная стенка, которая защищает протопласт от осмотического разрыва и придает клетке определенную форму; капсула (слизистый полисахаридный чехол вокруг клетки), цитоплазма и липиды.
biofile.ru
Клетка - дрожжи - Большая Энциклопедия Нефти и Газа, статья, страница 1
Клетка - дрожжи
Cтраница 1
Клетки дрожжей могут быть круглыми, продолговатыми, ли-монообразными или сильно удлиненными. [2]
Клетка дрожжей состоит из протоплазмы и оболочки. Оболочка клетки у некоторых дрожжей может в той или иной степени ослизняться, вследствие чего клетки склеиваются друг с другом и при развитии в жидких средах образуют быстро оседающие на дно сосуда хлопья. Оболочки клеток, которые не ослизняются, находятся во взвешенном состоянии. Дрожжи размножаются почкованием и лишь некоторые делением клетки пополам. [4]
Диаметр клеток дрожжей колеблется от 8 до 15 мкм. Форма их разнообразна: эллипсовидная, грушевидная, округлая, цилиндрическая. Размножаются вегетативным и половым путем. Вегетативные способы размножения - почкование и деление, половой способ размножения связан с образованием спор. При половом процессе слияние вегетативных клеток ведет к образованию сумок со спорами или сначала могут сформироваться споры, которые в последующем копулируют друг с другом. В каждой сумке образуется от 2 до 8, иногда 12 спор. Среди дрожжей есть аспорогенные, ложные дрожжи, неспособные к половому процессу и спорообразованию. Они относятся к классу несовершенных грибов. [5]
В клетках дрожжей и в растениях этот же процесс протекает в две стадии, через декарбоксилирование кетокислоты с образованием альдегида, который далее окисляется в жирную кислоту. Наличие в организме животных ферментов, могущих декарбокси-лировать кетокислоты в альдегиды без одновременного окисления, не доказано. [6]
При подсчете клеток дрожжей можно использовать краситель метиленовый синий. При этом в синий цвет окрашиваются только мертвые клетки. Живые клетки активно выводят краситель из клетки. Таким образом, можно подсчитать число жизнеспособных клеток, если учитывать только бесцветные или бледно-голубые клетки. [7]
Эргостерин в клетках дрожжей может быть в свободном виде и в виде эфиров жирных кислот. [8]
Что происходит с клетками дрожжей при нагревании и при растирании. [9]
Часто он накапливается в клетках дрожжей, бацилл. Для обогащения дрожжевых клеток гликогеном их выращивают на солодовой среде. [10]
Если сметана несвежая, иногда встречаются клетки дрожжей, молочной плесени - Geotrichum candidum ( Oidium lactis), споровых палочек. Для выделения молочнокислого стрептококка в чистую культуру используют сметану хорошего качества. [11]
По этому пути лизин образуется в клетках дрожжей, грибов, актиномицетов и некоторых видов водорослей. [12]
По мнению Пастера и его сторонников, цельная неповрежденная клетка дрожжей функционирует как организованный фермент брожения, якобы принципиально отличающийся от ряда содержащихся в различных клетках, в том числе и в дрожжах, неорганизованных ферментов - энзимов. [13]
По мнению Пастера и его сторонников, цельная, неповрежденная клетка дрожжей функционирует как организованный фермент брожения, принципиально отличающийся от ряда содержащихся в различных клетках, в том числе и в дрожжах, неорганизованных ферментов - энзимов. [14]
Разработана остроумная генетическая система, позволяющая заменять в клетках дрожжей нормальные гены на их модифицированные аналоги с помощью генно-инженерных манипуляций. В результате в клетке синтезируются измененные белки. Таким образом было показано, что гистоны Н2А и Н2В дрожжей можно лишить 10 - 30 концевых аминокислот и что это не влияет на сборку нуклеосом и структуру хроматина и вообще на жизнеспособность, клеток. Это особенно странно, если учесть высокую консервативность аминокислотных последовательностей гистонов. Возможно, N-концевые участки нуклеосомных гистонов необходимы не для сборки нуклеосом, а для другой цели, например для транспорта гистонов из цитоплазмы в ядро. [15]
Страницы: 1 2 3 4
www.ngpedia.ru
Винные дрожжи
Общая характеристика винных дрожжей
В образовании вина главную роль играют винные дрожжи, благодаря которым происходит спиртовое брожение углеводов.
Они представляют собой одноклеточные микроорганизмы, относящиеся к классу аскомицетов (сумчатых грибов). Дрожжи размножаются почкованием.
Дрожжевые клетки бывают яйцеобразной, эллипсоидной, овальной, булавоподобной формы, их длина составляет 5...12 мкм, ширина - 3...8 мкм.
Дрожжевая клетка состоит из оболочки, цитоплазматической мембраны, прилегающей к ней, цитоплазмы, рибосом и ядра (рис.).
Разрез дрожжевой клетки
1- аппарат Гольджи; 2 - рубец отпочкованой клетки; 3 - эндоплазматическая сетка; 4 - гранулы валютина; 5 - вакуоль; 6 - рибосомы; 7 - жировые капли; 8 - цитоплазматическая мембрана; 9 - маленькое ядро; 10 - митохондрия; 11 - клеточная оболочка; 12 - цитоплазма; 13 - ядро.
Оболочка винных дрожжей состоит из полисахаридов, белков, липидов и фосфатов. Оболочка пронизана порами, сквозь которые проходят к клетке питательные вещества, а также выводятся продукты распада.
Цитоплазматическая мембрана отделяет оболочку от цитоплазмы. Цитоплазматическая мембрана определяет осмотическое давление в клетке и обеспечивает выборочное передвижение питательных веществ из среды в клетку и вывод метаболитов из клетки. Состоит она из липидов и белка.
Перемещение веществ через цитоплазматическую мембрану проходит вследствие молекулярной диффузии и в результате активного движения, в котором принимают участие специфические ферменты.
Цитоплазма винных дрожжей состоит из воды, содержащей в растворимом состоянии углеводы, аминокислоты, минеральные соли и ферменты. Она имеет гетерогенную структуру и вяжущую консистенцию благодаря наличию белковых веществ. Цитоплазма молодых клеток гомогенная, а с возрастом появляются вакуоли, эндоплазматическая сетка, сосредоточение рибосом, жировые капельки, митохондрии.
Митохондрии – это дыхательный аппарат клетки, где сконцентрированы окислительные ферменты системы. Митохондрии имеют форму зернышек, палочек или нитей. Их химический состав представлен, в основном, белками и небольшим процентом липидов, полифосфатами, РНК и ДНК.
Питательные вещества, проникающие в клетку, адсорбируются и аккумулируются митохондриями и вследствие действия соответствующих ферментов подлежат быстрым превращениям. В митохондриях полностью осуществляется цикл трикарбоновых кислот и окислительное фосфорилирование, реакция активирования аминокислот в процессе биосинтеза белка, липидов и других соединений.
Рибосомы – это внутриклеточные частички, состоящие из РНК и белков и принимающие участие в биосинтезе белка.
Ядро является носителем потомственных свойств организма. В его состав входят молекулы ДНК и РНК, способствующие передаче наследственной информации, сохранению свойств микроорганизмов. При размножении клетки ядро разделяется на две части.
Обязательным органоидом дрожжевой клетки являются Вакуоли. Они представляют собой пустоты, наполненные клеточным соком, в состав которого входят белки, жиры, углеводы, минеральные вещества, ферменты, соли. Вакуоли являются своеобразным пищевым запасом клетки, используемым при голодании.
По внешнему виду клеток можно определять физиологическое состояние дрожжей. В дрожжевой разводке одновременно могут присутствовать молодые, зрелые, старые, почкующиеся клетки и мертвые. Наибольшую бродильную энергию имеют созревшие клетки.
В производстве вина встречаются, в основном, дрожжи рода Saccharomyces, пленочные и некоторые «дикие» дрожжи. К ним предъявляют ряд требований, среди которых, спиртоустойчивость (до 16,0 % об.) и спиртообразующая способность, высокая бродильная энергия, сульфито - и холодоустойчивость.
В своем развитии винные дрожжи проходят четыре основные фазы.
Лаг-фаза начинается с момента внесения разводки чистой культуры винных дрожжей в сусло – питательная среда. В этой фазе происходит процесс приспособления клеток к субстрату и не происходит их размножение, при этом размер клеток увеличивается.
Следующая фаза винных дрожжей – Логарифмическая – характеризуется увеличением популяции клеток и приростом биомассы, максимальной скоростью размножения дрожжей. В этой фазе повышается стойкость клеток к отрицательным внешним условиям и начинается спиртовое брожение.
Следующая стадия – Стационарная, при которой прекращается рост клеток дрожжей, а процесс спиртового брожения проходит интенсивно.
И последняя фаза развития винных дрожжей – фаза Затухания роста. Биомасса дрожжей снижается за счет интенсивного автолиза и использования дрожжами резервных веществ.
Виды винных дрожжей
Дрожжи, как возбудители спиртового брожения в виноделии, являются одноклеточными микроорганизмами. Их метаболизм основан на преобразовании одних веществ (сахаров) в другие (спирты, диоксид углерода, высшие спирты, сложные эфиры, альдегиды, диацетил и др.). Диаметр клеток дрожжей достигает 1...8 мкм, длина 2...12 мкм. Культуры дрожжей, которые выделили из винограда, и которые успешно. Сбраживают виноградное сусло, называют „винными дрожжами”.
Наиболее распространенными родами и видами винных дрожжей, которые играют главную роль в виноделии, являются Saccharomyces vini. Они активно размножаются и имеют высокую бродильную активность, а также быстро приспосабливаются к среде и определяют на конечном этапе состав вина. Количество спирта, который образуется при сбраживании 20 % мас. сахаров, составляет 11,6-12 %.
Дрожжи вида Saccharomyces oviformis практически полностью сбраживают сахара, которые содержатся в сусле и могут накапливать почти 18 % спирта. Разновидностью этого вида являются все хересные дрожжи и некоторые дрожжи шампанского производства, они образуют на поверхности вина пленку.
Широко распространены в виноделии такие виды винных дрожжей как Brettanomyces. Оптимальная температура их культивирования варьирует в пределах 31...32 оС. При сбраживании виноградного сусла они накапливают 11...12 % об. этилового спирта. Кроме того, вино обогащается летучими и не летучими кислотами. Благоприятной средой для размножения Brettanomyces в шампанском производстве является тиражный виноматериал.
Потомство одной клетки определенной расы, которая была отобрана путем селекции с учетом требований изготовления разных типов вин, представляет собой чистую культуру дрожжей.
Все расы винных дрожжей различаются между собой по активности сбраживания, скорости размножения, термо - и холодоустойчивости, кислотоустойчивости, сульфитоустойчивости, по способности к пенообразованию, скорости осветления виноматериалов после брожения, по способности к спиртообразованию, накоплению вторичных и побочных продуктов брожения, которые принимают участие в формировании аромата молодых вин.
В соответствии с требованиями биотехнологии и условиями изготовления виноматериалов, надо использовать такие расы дрожжей, которые имеют определенную особенность для данного типа вина. Так, для изготовления белых столовых и шампанских виноматериалов рекомендуют местные чистые культуры дрожжей, которые выделены в данном районе виноделия.
Особенного внимания заслуживает раса Феодосия I-19, которая активно сбраживает виноградное сусло до 24 % по сахаристости, имеет широкий температурный диапазон жизнедеятельности клеток. Эта культура сульфитостойкая, то есть сбраживает сусло с содержанием диоксида серы 200-250 мг/дм3.
С целью интенсификации брожения сусла необходимо, чтобы количество клеток чистой культуры дрожжей, которые вносятся, превышало количество клеток диких дрожжей, которые присутствуют в сусле.
На практике виноградное сусло подается на брожение нестерильным, после удовлетворительного отстаивания. При этом в каждом миллилитре сусла содержатся десятки тысяч клеток дрожжей вида Saccharomyces vini.
Биотехнология чистой культуры дрожжей характеризируется тем, что при внесении ее в нестерильное виноградное сусло, она противодействует диким дрожжам, которые размножаются быстрее, приспосабливаются к среде и успешно выжывают чистую культуру, которая была внесена.
Таким образом, применение данной формы брожения сусла в условиях практического виноделия не дает соответствующего эффекта, поэтому в большинстве винодельческих стран мира чистые культуры дрожжей, как правило, в первичном виноделии не нашли применения, так как в винодельческих районах жизнеспособные расы винных дрожжей были выделены естественным путем.
Внесение специальной чистой культуры дрожжей в нестерильное сусло обязательно только в случаях поступления на производство сусла с высокой титрируемой кислотностью, сахаристостью или пересульфитированное.
При низкой или высокой температуре брожения следует использовать холодоустойчивые или термостойкие расы винных дрожжей, поэтому на каждом заводе первичного виноделия к началу сезона переработки должен быть набор рас дрожжей.
Биотехнологические процессы при производстве столовых виноматериалов успешно проходят на препаратах активных сухих дрожжей. Это значительно снижает стоимость приготовления разводки в больших количествах, дает более глубокое выбраживание сахаров и повышает качество виноматериалов.
Добавить комментарий
vinograd-vino.ru
Клетка - дрожжи - Большая Энциклопедия Нефти и Газа, статья, страница 2
Клетка - дрожжи
Cтраница 2
Разработана остроумная генетическая система, позволяющая заменять в клетках дрожжей нормальные гены на их модифицированные аналоги с помощью генно-инженерных манипуляций - В результате в клетке синтезируются измененные белки. Таким образом было показано, что гистоны Н2А и Н2В дрожжей можно лишить 10 - 30 концевых аминокислот и что это не влияет на сборку нуклеосом и структуру хроматина и вообще на жизнеспособность клеток. Это особенно странно, если учесть высокую консервативность аминокислотных последовательностей гистонов. Возможно, N-концевые участки нуклеосомных гистонов необходимы не для сборки нуклеосом, а для другой цели, например для транспорта гистонов из цитоплазмы в ядро. [16]
Разработана остроумная генетическая система, позволяющая заменять в клетках дрожжей нормальные гены на их модифицированные аналоги с помощью генно-инженерных манипуляций. В результате в клетке синтезируются измененные белки. Таким образом было показано, что гистоны Н2А и Н2В дрожжей можно лишить 10 - 30 концевых аминокислот и что это не влияет на сборку нуклеосом и структуру хроматина и вообще на жизнеспособность клеток. Это особенно странно, если учесть высокую консервативность аминокислотных последовательностей гистонов. Возможло, N-концевые участки нуклеосомных гистонов необходимы не для сборки нуклеосом, а для другой цели, например для транспорта гистонов из цитоплазмы в ядро. [17]
Брожение - каталитический процесс; его вызывают образующиеся в клетках дрожжей вещества, относящиеся к классу ферментов, или энзимов, - биологических катализаторов белкового характера. Впоследствии оказалось, что активный дрожжевой сок содержит не один фермент, а сложную систему веществ белкового и небелкового характера, в которую входит несколько различных ферментов. При их участии превращение глюкозы в этиловый спирт протекает через ряд промежуточных соединений и является результатом нескольких реакций. Поэтому следует иметь в виду, что приведенное уравнение спиртового брожения выражает лишь окончательный результат процесса. [18]
Брожение - каталитический процесс; его вызывают образующиеся в клетках дрожжей вещества, относящиеся к классу энзимов, или ферментов - биологических катализаторов белкового характера. Впоследствии оказалось, что активный дрожжевой сок содержит не один фермент, а сложную систему веществ белкового и небелкового характера, в которую входит несколько различных ферментов. При их участии превращение глюкозы в этиловый спирт протекает через ряд промежуточных соединений и является результатом нескольких реакций. Поэтому следует иметь в виду, что приведенное уравнение спиртового брожения выражает лишь окончательный результат процесса. [19]
С целью получения этанола разработан метод, основанный на применении клеток дрожжей S. Биокатализатор на основе иммобилизованных клеток обладает высокой активностью, стабильностью и механической прочностью. Продуктивность по этанолу достигает 100 г / ( л ч), концентрация Сахаров на входе в реактор - 30 %, конверсия в этанол - 96 %, время полного сбраживания - 1ч, срок непрерывной эксплуатации биокатализатора достигал 10 - 12 мес. [20]
С целью получения этанола разработан метод, основанный на применении клеток дрожжей S. Биокатализатор на основе иммобилизованных клеток обладает высокой активностью, стабильностью и механической прочностью. Продуктивность по этанолу достигает 100 г / ( л ч), концентрация Сахаров на входе в реактор - 30 %, конверсия в этанол - 96 %, время полного сбраживания - 1ч, срок непрерывной эксплуатации биокатализатора достигал 10 - 12 мес. [21]
С Спирты, эфиры и слабые растворы щелочей растворяют липоидные вещества клеток дрожжей. Спирты даже в небольших концентрациях ( 3 - 4 %) тормозят почкование дрожжей. Размножение дрожжей при непрерывном брожении зависит главным образом от содержания питательных веществ, а не от содержания спирта в среде. [22]
Каньяр-Латур и одновременно и независимо Кютцинг ( 1838 г.) приписывали спиртовое брожение клеткам дрожжей, считавшихся живыми существами, по-видимому, растительного характера. В основе этих наблюдений лежит виталистическая теория брожения. Главный сторонник этой теории Пастер опубликовал в 1857 г. свое знаменитое исследование, посвященное спиртовому брожению, в котором он высказывает мысль, что брожение является процессом, непосредственно связ ишым с обменом дрожжевых клеток. Идя по пути старой теории Шталя, Либих ( 1839 г.) считал, наоборот, что брожение является разложением caxipa, обусловленным молекулярными колебаниями, вызванными химическими явлениями, происходящими в живых дрожжевых клетках. [23]
Биоценоз активного ила представлен большим разнообразием водорослей, отдельными нитями плесневых грибов и клетками дрожжей. [24]
В растворах серной кислоты концентрацией 0 35 - 0 6 % через 15 мин все клетки дрожжей сохраняют жизнеспособность; через 24 ч насчитывается лишь 2 % мертвых клеток. Молочнокислые бактерии в 0 15 % - ном растворе серной кислоты уже через 2 ч погибают, в 0 5 % - ном растворе в течение такого же времени погибают все бактерии. [25]
Большое значение при этом имеет правильная трансформация параметров процесса брожения: продолжительность пребывания сусла в аппаратах, численность клеток дрожжей, скорость притока свежего сусла и оттока зрелой бражки. [27]
В этом отношении интересна работа Н. Н. Постникова и В. В. Луцика, которые, исходя из модели плотности распределения клеток по возрасту, предлагают формулу, описывающую распределение клеток дрожжей по размерам [31], в которой учитываются переходы клеток в пределах трех возрастных групп - молодые клетки; почки и клетки, находящиеся на материнском теле; старые клетки. Учитывается также изменение размеров каждой группы и скорость роста в каждой группе. [28]
Особое значение цикла трикарбоновых кислот состоит в том, что он является связующим звеном между ферментными системами брожения и дыхания, являющимися главными источниками энергии для клеток дрожжей, и ферментными системами, обусловливающими синтез белковых веществ и других составных частей клеток. [29]
Зная существенную роль аденозинтрифосфорной кислоты, естественно возникает вопрос, откуда происходит АТФ, необходимая в больших количествах, но которая, как известно, содержится в клетках дрожжей в очень малой концентрации. [30]
Страницы: 1 2 3 4
www.ngpedia.ru
Дрожжи клеточный цикл - Справочник химика 21
| Рис. 13-1. Четыре последовательные фазы клеточного цикла типичной эукариотической клетки. После фазы М, которая состоит в делении ядра (митоз) и цитоплазмы (цитокинез) дочерние клетки вступают в интерфазу нового цикла. Интерфаза начинается с фазы С1, в которой возобновляются интенсивные биосинтетические процессы, резко замедленные во время митоза. Фаза 8-это период синтеза ДНК она заканчивается, когда содержание ДНК в ядре удвоится и хромосомы полностью реплицируются (теперь каждая хромосома состоит из двух идентичных сестринских хроматид ). Затем клетка вступает в фазу Сг, которая продолжается до начала митоза, т.е. фазы М. В фазе М удвоившиеся хромосомы конденсируются и становятся хорошо видимыми в световой микроскоп. Ядерная оболочка разрушается (исключение составляют одноклеточные эукариоты, например дрожжи, - у них она остается интактной) сестринские хроматиды расходятся и формируют два новых ядра, а цитоплазма делится с образованием двух дочерних клеток, имеющих по одному ядру. Процесс цитокинеза завершает фазу М, и начинается интерфаза следующего клеточного цикла. На рисунке представлен типичный 24-часовой цикл, однако длительность клеточного цикла у высших эукариот сильно варьирует она может быть короче 8 часов, а у взрослых животных - больше года, причем различия в основном зависят от |  |
| Рис. 13-19. Причинные связи межд некоторыми событиями клеточного цикла и их отношение к генам d у почкующихся дрожжей Полюсное тельце веретена (ПТВ) у дрожжей является эквивалентом центросомы. Старт означает точку бесповоротного вступления клетки в цикл деления и потерю возможности половой конъюгации (конъюгация может осуществляться только в фазе i). А.. Общая схема цикла стрелка, идущая от события а (или от событий а к б) к событию в, означает, что событие е не может произойти раньше события а (или аиб вместе). | 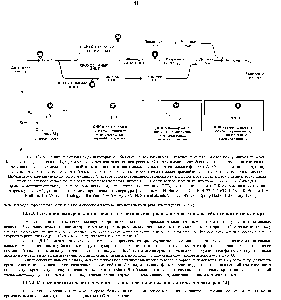 |
Скорость роста дрожжей зависит от свойств культуры и состава среды, а также от режима ферментации. Теоретически считают, что для полного развития клеточного цикла дрожжей необходимо 1—2 ч, но в производственных условиях развитие цикла идет 3—5 ч. [c.115]
Скорость роста простых свободноживущих организмов, таких как дрожжи, зависит главным образом от поступления питательных веществ. В условиях нехватки пищи дочерние клетки при быстрых циклах клеточного деления становились бы чрезвычайно мелкими поэтому клеткам необходим механизм, регулирующий скорость прохождения клеточного цикла и, в частности, хромосомного цикла в соответствии со скоростью роста клетки (рис. 13-20) Как осуществляется такая регуляция [c.411]
Сравнение рис. 1, а, б показывает, что в структуре митотического цнкла почкующихся и делящихся дрожжей имеются значительные различия. Главное различие заключается в положении S-фазы, которая у почкующихся дрожжей находится в начале, а у делящихся — в конце клеточного цикла. Сходным является то, что как почкующиеся, так и делящиеся дрож- [c.7]
Фибробласты (такие, как различные типы мышиных клеток ЗТЗ) обычно делятся быстрее, если расположить их в культуральной чашке не слишком плотно и использовать культуральную среду, богатую питательными веществами и содержащую сыворотку - жидкость, получаемую при свертывании крови и очищенную от нерастворимых сгустков и кровяных клеток. При нехватке каких-либо важных питательных веществ, например аминокислот, или при добавлении в среду ингибитора белкового синтеза клетки начинают вести себя примерно так же, как описанные выше дрожжевые клетки при недостатке питания средняя продолжительность фазы Gt возрастает, но на остальной части клеточного цикла все это почти не сказывается. Как только клетка прошла через Gi, она уже неизбежно и без задержки проходит фазы S, Сг и М независимо от условий среды. Эту точку перехода в поздней фазе Gi часто называют точкой рестрикции (R), потому что именно здесь клеточный цикл еще может приостановиться, если внешние условия препятствуют его продолжению Точка рестрикции соответствует точке старта в клеточном цикле дрожжей так же как и дрожжей, она может отчасти служить механизмом, регулирующим размеры клетки. Однако у высших эукариот ее функция более сложна, чем у дрожжей, и в фазе G может быть несколько слегка различающихся точек рестрикции, связанных с различными механизмами контроля клеточной пролиферации. [c.416]
Исследования на мутантных дрожжевых клетках показали, что для начала синтеза ДНК необходимо удвоение структуры, находящейся на ядерной оболочке,-полярного тельца веретена. Аналогом этой структуры в животной клетке является центриоль, которая действует и как часть важного центра организации микротрубочек, тесно связанного с интерфазным ядром (клеточный центр, см. разд. 10.4.2), и как компонент каждого из полюсов веретена во время митоза. Центриоль удваивается, по-видимому, путем матричного процесса один раз за клеточный цикл (см. рис. 11-19 и разд. 10.4.4). Возможно, что достижение определенной стадии в процессе ее удвоения (так же как и в случае полярного тельца веретена у дрожжей) представляет собой критический момент в цепи событий, инициирующих репликацию ДНК. К сожалению, пока нет возможности блокировать удвоение центриоли и проверить таким образом это предположение. [c.158]
Один из главных выводов, к которому привели такого рода исследования, состоит в том, что у дрожжей имеется много различных белков, участвующих в том или ином событии, связанном с клеточным циклом. Эти белки не обязательно должны синтезироваться только на той стадии цикла, на которой они работают, поскольку после инъекции нормальной цитоплазмы большинство мутантных клеток способно пройти несколько нормальных делений при высокой температуре. Это означает, что у дрожжей на протяжении всего цикла необходимых белков намного больше, чем нужно для одного цикла. По аналогии можно полагать, что и в клетках млекопитающих многие белки, синтезируемые все время с постоянной скоростью, могут функционировать лишь на определенных стадиях цикла (см. рис. 11-31). [c.171]Мутации, специфически влияющие на отдельные компоненты механизма клеточного цикла, нельзя обнаружить по одной лишь утрате способности мутантных клеток к делению, так как к этому будет приводить любой летальный дефект. Мутации цикла клеточного деления Ыс-се11-ё1у181оп-еуе1е) более достоверно выявляются по тому, как они блокируют или нарушают специфическую фазу клеточного цикла при пермиссивной гемпературе (рис. 13-18). У почкующихся дрожжей наличие и размеры почки служат простым визуальным индикатором, показывающим, какой )тап клеточного цикла блокирован у данного мутанта с(1с в случае делящихся дрожжей нужны более сложные подходы с использованием методов анализа клеточного цикла, о которых говорилось выше. [c.410]
При исследовании мутантных дрожжей были также получены важные сведения о биохимических процессах, связанных с клеточным циклом. Возник, например, вопрос, в скольких различных точках цикла действуют белки, идентифицированные с помощью мутаций d . Много разнообразных белков требуется, в частности, для репликации ДНК. Большое число различных продуктов, кодируемых генами d , должно работать одновременно для завершения фазы S так оно и оказалось в действительности. Аналогичным образом было показано, что белки у дрожжей распадаются по крайней мере на семь групп, каждая из которых участвует в одном из семи биохимических событий, необходимых для завершения цикла. Удалось определить последовательность, в которой действуют эти семь групп, и бьш поставлен вопрос необходимо ли завершение одного биохимического события из данной серии, для того чтобы мог начать работать какой-либо белок из следующего набора [c.171]
Таким образом, первая группа фактов побуждает искать причину рака в действии генетического материала, вносимого в клетку извне, а вторая — искать генетические причины рака в самой клетке. Эти подходы объединяют сведения о том, что в нормальных клетках существуют так называемые протоонкогены — гены, гомологичные онкогенам ретровирусов. Протоонкогены чрезвычайно консервативны и сходны в геномах человека, мыши, дрозофилы и даже дрожжей. Некоторые из них контролируют нормальное протекание клеточного цикла. Нельзя сказать, что механизм канцерогенеза выяснен, однако наиболее вероятной причиной представляется злокачественная трансформация клетки вследствие нарушения экспрессии некоторых ее генов (онкогенов, протоонкогенов) в результате мутационных или модификационных изменений, а также в результате вирусной инфекции. [c.540]
Помимо химических средств, гипотермии и гипоксии модификация радиорезистентности производилась с помощью фракционированного облучения в невысоких летальных дозах. На клетках асцитной карциномы было показано, что при увеличении времени между двукратным облучением одновременно с возрастанием радиорезистентности объектов общее содержание эндогенных тиолов увеличивается. Параллельное исследование радиорезистентности и уровня сульфгидрильных групп проводилось также на клетках, находящихся на разных стадиях роста и клеточного цикла. Так, Э. Я. Граевский (1969) привел сравнительные данные из работ по изучению изменения тиолов и радиорезистентности микроспор в процессе клеточного деления. Оказалось, что в процессе мейоза и митоза происходят однонаправленные изменения содержания тиолов в клетках и их устойчивости (устойчивость оценивалась по выходу хромосомных аберраций) к действию ионизирующей радиации. Динамика изменения уровня эндогенных сульфгидрильных групп в зависимости от изменения радиорезистентности прослежена также на синхронно делящейся икре морских ежей в различных стадиях клеточного цикла, на растущих клетках асцитной карциномы Эрлиха в процессе ее старения, на синхронной культуре клеток разных штаммов хлореллы в процессе клеточного деления, на клетках в различных фазах роста. Эти данные позволили авторам заключить, что изменения радиочувствительности в цикле связаны не только с изменением генетического аппарата в клетке, но и с варьированием содержания внутриклеточных тиолов, выполняющих функции эндогенных радиопротекторов. Эти представления получили дополнительное обоснование в работе Ю. В. Корогодиной и др. (1975). Так, на диплоидных дрожжах (штамм Мегри 139 В) было установлено, что клетки, находящиеся в логарифмической фазе роста, в отличие от стационарной фазы более радиорезистентны и содержат в полтора раза больше сульфгидрильных групп. Авторы считают, что именно высокий уровень тиолов почкующихся дрожжевых клеток может определять их повышенную радиорезистентность. [c.283]
А. Дрожжи важны для исследований клеточного цикла в первую [c.241]
Б. Наиболее существенные регуляторы клеточного цикла у дрожжей-это тип питания и пол. [c.241]
Синтез ДНК, как мы уже выше видели, осуществляется лишь в течеппе короткого периода клеточного цикла, синтезы РНК и белка, по данным многих авторов, как у почкующихся дрожжей (Halvorson et al., 1964 и др.), так и у делящихся дрожжей (Wain, 1971 и др.) протекают [c.16]
Такие сравнения белков важны еще и в том отношении, что сходные структуры часто предполагают и сходные функции. Можно избежать многолетних экспериментальных исследований, установив гомологию аминокислотной последовательности с белком, функция которого известна. Например, гакие гомологии последовательностей впервые указали на то, что некоторые регуляторные гены клеточного цикла дрожжей и некоторые гены, вызывающие раковое перерождение клеток млекопитающих, кодируют протеинкиназы. Таким же способом было определено, что многие из белков, контролирующих морфогенез у плодовой мушки Drosophila, являются белками регуляторного гена, а один белок, участвующий в морфогенезе, был идентифицирован как сериновая протеиназа. [c.150]
Репродуктивный цикл типичной эукариотической клетки можно подразделить на четыре фазы, обозначаемые Gi (от конца митоза до начала синтеза ДНК), S (синтез ДНК), G2 (от конца синтеза ДНК до начала митоза) и М (митоз). Каждая S- и М-фаза инициируется растворимым цитоплазматическим фактором [активатором S-фазы и М-стимулирующим фактором (MPF ) соответственно]. Активатор S-фазы образуется на протяжении всей S-фазы и может также действовать как фактор, задерживающий подготовку к М-фазе до тех пор, пока не завершится репликация ДНК. М-стимулирующий фактор может быть обнаружен в М-фазных клетках многих организмов - от дрожжей до млекопитающих, и его активность, возможно, регулируется фосфорилированием. В яйцах, быстро подвергающихся дроблению, таких как у Хепорш, клеточный цикл укорочен и упрощен. В этом случае цикл, по-видимому, регулируется взаимосвязанными колебаниями активности MPF и концентрации циклина. [c.407]
Дрожжи являются одноклеточными грибами и составляют большую группу довольно разнородных организмов. Поскольку они размножаются почти так же быстро, как бактерии, и размеры их генома меньше 1/1000 генома млекопитающих, они оказались чрезвычайно полезными для генетического анализа клеточной биологии эукариот. Хотя яйца Хепорш-шкшочшсльно ценный объект для изучения биохимических и цитофизиологических аспектов регуляции клеточного цикла, для генетических исследований этот объект неудобен. Напротив, работа с дрож- [c.407]
Если повысить температуру до рестриктивного уровня, у большинства мутантов сс1с клеточный цикл останавливается на той стадии, на которой действует продукт гена сб/с. Как правило, клетка теряет способность переходить к следующей стадии цикла, и это означает, что начало каждого процесса находится в зависимости от завершения предыдущего процесса Таким образом, у дрожжей, как и у млекопитающих, большинство этапов клеточного цикла, но-видимому, связаны между собой как звенья единой цепи. Эта связь была более тщательно проанализирована в экспериментах с клетками, содержащими разные комбинации различных мутаций сс1с. Как показали результаты, события хромосомного цикла образуют ряд зависимых друх от друга этапов, который не связан жестко с событиями цитоплазматического цикла (рис. 13-19). Например, хотя цитокинеза не произойдет, если предотвратить деление ядра, тем не менее мутанты сс1с, не способные пройти цитокинез из-за дефектов в механизме формирования почки, все же осуществляют повторные циклы синтеза ДНК и деления ядра. Но-видимому, общим правилом не только для дрожжей, но и для клеток млекопитающих, насекомых и многих других организмов является то, что хромосомный цикл может продолжаться, даже если цитокинез предотвращен. В самом [c.410]
Рассмотрим пока лишь события, происходящие в ядре. Здесь мы находим строгую последовательность реакций, специфичных для определенных стадий клеточного цикла (рис. 11-33). У дрожжей эта последовательность организована подобно линейной головоломке, в которой каждый предыдущий фрагмент должен занять свое место, прежде чем можно будет уложить последующий. В результате все события происходят в строгом циклическом порядке. Например, репликация ДНК может начаться только после удвоения полярного тельца веретена деление ядра-только после репликации ДНК следующая дупликация полярного тельца-только после разделения ядра. В принципе ход организованного таким образом цикла может контролироваться просто скоростью добавления предобразованных кусочков головоломки , а синтез белков, специфичных для данной стадии цикла, может в этот момент и не требоваться. [c.171]
Интересно, что модель сходного типа была предложена и для объяснения регуляции клеточного цикла у более примитивных организмов. Однако у некоторых из них, например у S hizosa haromy es ротЬе (дрожжи), амебы (простейшие) и Physarum (миксомицеты), фаза Gi вообще отсутствует, и после митоза клетки прямо вступают в фазу S. У этих клеток точка рестрикции, по-видимому, контролирующая клеточное деление, находится в начале фазы М. [c.173]
Генетические исследования на мутантах дрожжей показали, что, хотя растущие клетки удваивают свое содержимое в целом независимо от фаз цикла, в процессе их роста осуществляется также ряд последовательных процессов, специфически связанных с определенными фазами клеточного цикла. Многие из этих процессов, по-видимому, включают построение надмолекулярных структур из предсуществующих молекул. Завершение сборки этих структур делает возможной следующую стадию сборки. Про.чежуток между последовательными митозами определяется суммарным временем, необходимым для осуществления всех звеньев такой цепи событий. Одна из реакций в этом каскаде может служить специальным регулятором, обеспечивающим нормальные размеры клеток. В клетках млекопитающих подобная регуляторная система препятствует прохождению точки рестрикции в фшзе (а значит, и началу синтеза ДНК) до тех пор, пока не будут синтезированы все ко.мпо-ненты, необходимые для завершения всех последовательных процессов сборки в предстоящих фазах 8, 61 и М. [c.175]
Высокоспецифичные протеиназы играют важную роль в регуляции клеточного цикла, в частности при переходе от вегетативного роста к образованию спор у дрожжей и бактерий. У мутантов дрожжей с низким уровнем протеиназ А и В снижена способность к споруляции. Выделен температурно-чувствительный мутант по споруляции Вас. subtilis, у которого этот дефект связан с повреждением структурного гена для внутриклеточной протеиназы. У Вас. thuringiensis с началом спорообразования специфическая протеиназа расщепляет -субъединицу РНК-полимеразы, осуществляя необходимую модификацию фермента. [c.56]
Митохондрии — органеллы, окруженные мембраной. Они содержат ДНК, РНК и широкий спектр ферментов, необходимых для окисления уксусной кислоты до СОг, восстаповления кислорода до воды и использования избытка возникаюгцей при этом энергии для.образования богатых энергией фосфатных связей. Конечный продукт аденозиптрифосфат (АТФ) — это энергетический банк клетки, из него клетка по мере необходимости черпает энергию для всех реакций, протекающих с затратой энергии. Хлоропласты — не менее сложные структуры, в состав которых входят ДНК, РНК, весь фотосинтетический аппарат с фоторецепторами, абсорбирующими солнечный свет, и ферментами для образования глюкозы из СОг и Ог. Обе эти органеллы самореплицируются, т. е. в клеточном цикле число их также удваивается и они распределяются между дочерними клетками. Было обнаружено, что у дрожжей по каким-либо случайным причинам почка может не получить митохондрий от материнской клетки. Эта потеря невосполнима. Родительские клетки дрожжей, содержащие митохондрии, относительно крупные с ненарушенной функцией дыхания (в присутствии кислорода они окисляют глюкозу до СОг через уксусную кислоту). У повой разновидности дрожжей, возникшей из несчастливой почки, нет митохондрий, клетки мелкие, и растут они очень медленно, используя ферментативное окисление глюкозы до СОг и этилового спирта этот процесс не требует кислорода. Родительская клетка использует его только в анаэробных условиях. [c.69]
Мутанты, т. е. организмы или клетки, несущие мутацию, у которых на разных стадиях блокирован клеточный цикл, получены у ряда одноклеточных организмов — дрожжей родов Sa haromy es и S hizosa haromy es, водоросли hlamidomonas, плесневого фиба [c.62]
Таким образом, первоначальная попытка идентифицировать гены дрожжей, отвечающие за высокую спонтанную г/го--мутабильность, привела нас к получению мутаций в ядерных генах, которые, по всей вероятности, занимают узловые позиции в системе координированной регуляции генетической стабильности, смены фаз клеточного цикла и радиочувствительности. Современные достижения геномики существенно облегчили идентификацию этих генов. Представляются целесообразными как дальнейший детальный анализ уже обнаруженной регуляторной роли генов SRM, так и выявление и исследование функций этих генов применительно к таким, например, существенным аспектам генетической регуляции, как метаболическая индукция/репрессия и сайленсинг. [c.103]
Д. В клеточном цикле дрожжей существует критическая точка, называемая , прохождение которой означает, что цикл клеточного деления будет обязательно завершен. [c.241]
chem21.info




















 |  |  |  |  |  |  |  |  |  |  |  |  |  |  |
 Пример видео 3 Пример видео 3 |  Пример видео 2 Пример видео 2 |  Пример видео 6 Пример видео 6 |  Пример видео 1 Пример видео 1 |  Пример видео 5 Пример видео 5 |  Пример видео 4 Пример видео 4 |
Администрация муниципального образования «Городское поселение – г.Осташков»