



Базидиомицетовые дрожжи. Базидиомицеты дрожжи
Базидиомицетовые дрожжи
В отличие от аскоспоровых дрожжей, все дрожжи, которые относят к базидиомицетам, образуют в ходе полового размножения специальные клетки - базидии, несущие экзогенные споры (базидиоспоры). Значительную часть жизненного цикла занимает дикариотическая фаза, представленная дикариотическим мицелием с пряжками. Большинство видов гетероталличны и поэтому многие из них долгое время были известны только в гаплофазе как несовершенные дрожжи.
Жизненные циклы базидиомицетовых дрожжей также достаточно разнообразны. Активное исследование жизненных циклов базидиомицетовых дрожжей началось лишь в конце 60-х - начале 70-х годов XX в., после того как японский миколог Исао Банно опубликовал результаты открытия полового цикла у хорошо известных и ранее считавшихся несовершенными красных дрожжей Rhodotorula glutinis. Ему удалось подобрать типы спаривания среди большого числа проверенных штаммов, и в результате их скрещивания он наблюдал образование дикариотического мицелия, на котором формировались структуры, морфологически сходные с хламидоспорами, то есть крупные клетки с сильно утолщенными оболочками и большим запасом внутриклеточных липидов. Именно в них затем происходило слияние двух ядер и последующий мейоз с восстановлением гаплоидного состояния. Эти клетки получили название телиоспор.
При прорастании они образовывали промицелий с поперечными септами, делящими его на 4 клетки. Эти клетки отпочковывали гаплоидные споридии. Совершенная стадия Rhodotorula glutinis была названа Банно Rhodosporidium toruloides. Позже сходный цикл был описан для некоторых баллистоспоровых дрожжей и для нескольких видов несовершенного рода Candida. Их телиоспоровые стадии получили соответственно родовые названия Sporidiobolus и Leucosporidium.
 Сейчас похожие полные или не завершенные жизненные циклы известны для многих базидиомицетовых дрожжей. Они различаются формой телиоспор, характером их прорастания и морфологией базидий. Другой тип жизненного цикла базидиомицетов с дрожжевой анаморфой известен для дрожалковых грибов порядка Tremellales.
Сейчас похожие полные или не завершенные жизненные циклы известны для многих базидиомицетовых дрожжей. Они различаются формой телиоспор, характером их прорастания и морфологией базидий. Другой тип жизненного цикла базидиомицетов с дрожжевой анаморфой известен для дрожалковых грибов порядка Tremellales.
 Фото: eelke dekker
Фото: eelke dekker
В культурах отдельные базидиоспоры гетероталличных видов дают начало дрожжевым стадиям. Мицелий обычно развивается только при смешивании штаммов совместимых типов спаривания. Смешение соответствующих типов спаривания приводит к быстрой реакции клеток на выделяющиеся феромоны; почкование прекращается, развиваются конъюгационные трубки, через которые клетки копулируют, происходит плазмогамия и развивается мицелиальная дикариотическая стадия. У многих видов в подходящих условиях вскоре после формирования дикариотического мицелия начинается развитие плодовых тел. В плодовых телах образуются 2-4-клеточные гетеробазидии. Освободившиеся базидиоспоры могут размножаться вегетативно почкованием или формированием баллистоспор.
Характеристики жизненного цикла играют большую роль в систематике базидиомицетовых дрожжей. Однако очень часто у базидиомицетовых дрожжей, изолируемых из природных местообитаний, не удается наблюдать полный жизненный цикл при культивировании на лабораторных средах, поэтому их идентификация и классификация вызывают большие трудности. Значительные успехи в систематике таких анаморфных базидиомицетовых дрожжей достигнуты в последние десятилетия, особенно в связи с внедрением в таксономическую практику молекулярно-биологических методов. Как оказалось, группирование базидиомицетовых дрожжей на основе молекулярно-биологических признаков, в частности нуклеотидных последовательностей рРНК, часто не совпадает с характеристиками их жизненного цикла, особенно такими, как способность к образованию телиоспор, морфология базидий. Поэтому классификация базидиомицетовых дрожжей еще далека от завершенности и требует дальнейших исследований.
biofile.ru
Базидиомицетовые дрожжи
Почти все грибы, обозначаемые термином «базидиомицетовые дрожжи» - это диморфныеорганизмы, имеющие мицелиальную телеоморфную и одноклеточную почкующуюся анаморфную стадии. Одна из главных задач их классификации - выяснение филогенетических связей между базидиомицетовыми дрожжами и «истинными» мицелиальными базидиомицетами. Современная классификация диморфных базидиомицетов в основном базируется на анализахнуклеотидных последовательностейрДНК. В классификации базидиомицетовых дрожжей большую роль играют такие признаки, какмоносахаридный составклеточных стенок,морфология базидий,ультраструктура септмицелия, образованиекрахмалоподобных соединений, способность к ассимиляцииинозита. Как оказалось, именно эти признаки в наибольшей степени отражают филогенетическое родство видов и достаточно хорошо коррелируют с группированием базидиомицетовых дрожжей на основании нуклеотидных последовательностей. Ультраструктурные, биохимические и молекулярно-биологические исследования, проведенные в последние десятилетия, показали, что базидиомицетовые дрожжи не являются монофилетической группой и распределены по трем основным филогенетическим линиям базидиомицетов, которые выделяются в качестветрех классов: Hymenomycetes, Urediniomycetes и Ustilaginomycetes.
Роды базидиомицетовых дрожжей
| Класс Urediniomycetes
| |||||||||||||||||||||||||||||
|
| Класс Ustilaginomycetes
| ||||||||||||||||||||||||||||
|
| Класс Hymenomycetes
| ||||||||||||||||||||||||||||
|
| Анаморфные роды базидиомицетового аффинитета
|
Литература
Бабьева И.П. Дрожжи в биогеоценозах разных природных зон. В кн.: «Почвенные организмы как компоненты биогеоценоза», М.: Наука, 1984
Бабьева И.П. Дрожжи. В кн.: «Мир растений», М.: Просвещение, 1990, т. 2
Бабьева И.П., Голубев В.И. Методы выделения и идентификации дрожжей. М.: Пищевая промышленность, 1979
Бабьева И.П., Горин С.Е. Почвенные дрожжи. М.: Изд‑во МГУ, 1987
Берри Д. Биология дрожжей. М.: Мир, 1985
Квасников Е.И., Щелокова И.Ф. Дрожжи. Биология. Пути использования. Киев: Наукова Думка, 1991
Кудрявцев В.И. Систематика дрожжей. М.: Изд‑во АН СССР, 1954
Новое с систематике и номенклатуре грибов. Ред. Ю.Т.Дьяков, Ю.В.Сергеев. М.: Национальная академия микологии. Медицина для всех, 2003, 494 с.
Barnett J.A., Payne R.W., Yarrow D. A guide to identifying and classifying yeasts. Cambridge: Cambridge Univ. Press, 1991
Biology and activity of yeasts. Eds. F.A.Skinner, S.M.Passmore, R.R.Davenport. London et al., 1980, 310 p.
The expanding realm of yeast-like fungi. Eds. G.S. de Hoog, M.Th.Smith, A.C.M.Weijman. Amsterdam: Elsevier Science Publ., 1987, 510 p.
The yeasts. A taxononic study. Second ed. Ed. J.Lodder. Amsterdam: North-Holl. Publ. Co., 1970, 1386 p.
The yeasts. A taxonomic study. Ed. N.J.W.Kreger‑van Rij. Third edition. Amsterdam: Elsevier, 1984, 1082 p.
The yeasts. A taxonomic study. Eds. C.P.Kurtzman, J.W.Fell. Fourth revised and enlarged edition. Amsterdam: Elsevier Science B.V., 1998, 1055 p.
Phaff H.J., Miller M.W., Mrak E.M. The life of yeasts. 2‑d edition. Cambridge: Harvard Univ. Press, 1978.
The Yeasts. Eds. A.H.Rose, J.S.Harrison. Biology of yeasts. London et al.,: Acad. Press, vol. 1‑3, 1987.
studfiles.net
Базидиомицеты - Справочник химика 21
Базидиомицеты считаются наиболее высокоразвитой группой грибов. Характерный для них орган спороношения — базидия — по своему расположению (это клетка на конце гифы) соответствует аску. От бази-дии отшнуровываются, как правило, четыре одноядерные гаплоидные базидиоспоры. Подобно аскоспорам, они образуются в результате плазмогамии, кариогамии и мейоза. Два последних процесса протекают соответственно в аске или в базидии. [c.72] Нельзя, однако, говорить о полном отсутствии полового процесса у несовершенных грибов. У них, так же, как у базидиомицетов или у других аскомицетов, доказано наличие парасексуального процесса. Плазмогамия, кариогамия и мейоз имеют место и у них, но не в определенных участках вегетативного тела и не на определенных стадиях развития. В норме первичный мицелий несовершенных грибов бывает гомокариотическим, т. е. содержит ядра только одного типа. В результате объединения протопластов, содержащих ядра разного типа, возникают гетерокарионы. Введенное в мицелий чужое ядро размножается, и образовавшиеся дочерние ядра распространяются по мицелию. Время от времени происходят кариогамия и мейоз. Таким образом, парасексуаль-ный цикл обеспечивает примерно столь же эффективную рекомбинацию ядерного материала, как и истинный половой процесс. [c.75]Для многих базидиомицетов характерны пряжки (рис 3), образующиеся на мицелии и участвующие в синхронном процессе деления ядер, а в мицелиальных септах — долипоры (см рис 32) [c.34]
К числу аскомицетов относятся очень ценные съедобные трюфели Ц сморчки. Однако большинство съедобных грибов представляют собой плодовые тела базидиомицетов. К базидиомицетам относятся также, ржавчинные грибы, наносящие колоссальный ущерб урожаю пшеницЫ и других зерновых культур. [c.47]
| Рис. 46.2. Средняя токсичность для дереворазрушающих базидиомицетов после вымачивания |  |
Дрожжи — это грибы, приспособившиеся к существованию в среде с высоким содержанием сахара они остаются обычно одноклеточными и размножаются путем почкования (рис. 1-8). Время от времени их гаплоидные клетки попарно сливаются и образуют диплоидные клетка и половые споры. Одни дрожжи относятся к аскомицетам, другие — к базидиомицетам. Sa haromy es erevisiae, активное начало как пекарских, так и пивных дрожжей, является аскомицетом, способным к неог-. раниченному росту как в диплоидной, так и гаплоидной фазе, причец диплоидные клетки несколько крупнее гаплоидных [37]. [c.47]
IV. Базидиомицеты (Basidiomy etes) —базидиальные грибы, имеющие многоклеточный. мицелий, размножаются в основном половым путем. [c.501]
Из плодового тела, сформированного грибом Аиг1са1аг1а зр., относящимся к классу базидиомицетов, выделен ацетилироваииый (1—>-3)-а-Л-маннан, обладающий значительным антпвоспали-тельным действием [91]. [c.266]
Изопропоксианилид 2-(трифторметил) бензойной кислоты (12) (флутранил, монкут) — твердое вещество, т. пл. 108°С. Малорастворим в воде, хорошо растворяется в органических растворителях, ЛД50 для экспериментальных животных более 10000 мг/кг. Активный фунгицид против базидиомицетов, норма расхода 0,3—1 кг/га. [c.204]
Аскомицеты вместе с базидиомицетами относятся к высшим грибам (эумицетам). Для последних характерны гифы, разделенные поперечными перегородками, и образование конидиоспор. Клетки со жгутиками у высших грибов не встречаются. [c.65]
Цикл развития гименомицетов. Из проросшей базидиоспоры образуется первичный мицелий, состоящий из гиф, разделенных перегородками на одноядерные клетки. Вторичный, дикариофитный, мицелий возникает в тех случаях, когда встречаются гифы двух совместимых штаммов и их одноядерные протопласты сливаются (плазмогамия). При каждом клеточном делении происходит сопряженное деление обоих ядер. У многих базидиомицетов деление клеток и ядер сопровождается образованием так называемой пряжки (рис. 1.35). Такой механизм обеспечивает получение новой клеткой по одному дочернему ядру каждого типа. У клетки, готовой к делению, между ядрами аиЬ образуется нечто вроде крючка, который загибается назад переднее ядро Ь переходит в этот крючок (пряжку), после чего оба ядра делятся. Затем передняя часть клетки, содержащая дочерние ядра а и Ь, отделяется от задней части поперечной перегородкой. Одновременно пряжка сливается с исходной клеткой, в которую при этом возвращается ядро Ь. Передняя [c.72]
КЛАСС ВА8ЮЮМ СЕТЕ8. Большинству людей на земле базидиомицеты известны по плодовым телам съедобных н несъедобных шляпных грибов (белый гриб, груздь, мухомор и т. д.), в изобилии вырастающих в лесных зонах различных географических широт земною шара. Съедобные шляпные грибы составляют существенное звено в питании многих народов. Поэтому вполне резонна постановка задачи выращивать в промышленных условиях биомассу тех или иных съедобных грибов с присущими им ароматом и вкусом. [c.199]
Секреторные (выделительные) ткани в виде специализированных морфологических структур отсутствуют у грибов, хотя экзоцитоз (см ) присущ клеточной мембране и по этой функции она Зшодобляется секреторной ткани Из клеток грибов секретируются многие первичные и вторичные метаболиты (ферментные и неферментные белки, пигменты и другие вещества) Структуры, подобные млечникам у растений, имеются у некоторых базидиомицетов (скрипица, рыжик) В этих случаях между гифами и окружающими их клетками образуются ситовидные пластинки и гифы становятся как бы млечными трубками, содержашдми млечный сок (эмульсия смол, масел, пигментов, горьких и других веществ в воде) [c.118]
Облегченная диффузия осуществляется при участии мембранных пор, генерирующих ион-проводящие пути, ионофоров-пепти-дов, формирующих ионные каналы (например, декалептиды— антаманид у базидиомицетов, грамицидин С у некоторых бацилл и др ), интегральных и периферических мембранных белков, действующих взаимосвязанно (например, пермеазы и др ) [c.267]
В качестве гидролизующих агентов используют кислоты или, реже, ферментные комплексы. В любом случае матричный лигнин остается негидролизованным и от него стремятся избавиться предварительной делигнификацией целлюлозных материалов различными способами, в том числе — паракрекингом, или обработкой водяным паром под давлением 7—8 ати. Можно использовать биологический способ делигнификации, когда на целлюлозосодержащем материале выращивают дереворазрушающие грибы из класса базидиомицетов. Эти последние делают целлюлозный материал доступным в качестве корма для жвачных животных (содержание лигнина в нем не должно превышать 18—20%). [c.521]
К базидиальным принадлежат грибы, плодовые тела которых общеизвестны как съедобные и ядовитые. Они, как и другие базидиомицеты и архимицеты, ие входят в сферу интересов микробиологии. [c.73]
Природные ацетиленовые углеводороды. — До 1950 г. очень небольшое количество ацетиленовых соединений было обнаружено в природных объектах. Исследования Сёренсена в Норвегии, Больмана в Германии и Джонса и Уайтинга в Англии показали, что ацетиленовые кислоты, спирты, кетоны и углеводороды часто встречаются, хотя и в малых количествах, в высших грибках (базидиомицетах) и высших растениях (сложноцветных и зонтичных). Типы углеводородов, выделенных Сёренсеном из сложноцветных растений, представлены формулами, установленными для ряда С1з-углеводородов [c.279]
ИЗ нее, упаковочных и других материалов от повреждений микрогрибами [2]. Достаточно э( )фективен фурметамид в отношении дереворазрушающ,их базидиомицетов (рис. 46.1, 46.2), менее токсичны трибутилоловооксид и пентахлорфенол [1]. [c.499]
Биохимия Том 3 (1980) -- [ c.47 ]Очерк химии природных соединений (2000) -- [ c.144 ]
Общая микробиология (1987) -- [ c.51 , c.159 , c.173 , c.174 , c.344 ]
Некоторые вопросы химии серусодержащих органических соединений (1963) -- [ c.86 ]
Биотехнология - принципы и применение (1988) -- [ c.301 , c.364 ]
Малый практикум по низшим растениям (1994) -- [ c.137 , c.139 , c.156 , c.208 , c.209 , c.235 , c.246 ]
Микробиология (2003) -- [ c.33 , c.35 ]
chem21.info
Гапло-диплоидные дрожжи
Существуют дрожжи, у которых вегетативное размножение может происходить как в гаплоидной, так и в диплоидной фазах. Длительность той или другой фазы зависит от вида и от условий роста. Среди этих дрожжей есть как гомоталличные, так и гетероталличные виды.
Saccharomyces cerevisiae. Эти дрожжи вегетируют преимущественно в диплоидном состоянии, но у них имеется короткая вегетативная гаплоидная фаза. Диплоидная клетка в условиях дефицита легкодоступных источников углерода прекращает почковаться, и ядро ее делится мейотически. В результате она превращается в аск с 4 гаплоидными аскоспорами, которые после освобождения из аска прорастают и образуют гаплоидное поколение. Гаплоидные клетки обычно мельче диплоидных и имеют более округлую форму. Шрамы почкования у них сближены, почки образуются группами в одном локусе. После нескольких циклов почкования две клетки конъюгируют и сливаются, восстанавливая диплоидное состояние.
Metschnikowia pulcherrima.Диплоидные клетки этого вида размножаются вегетативно почкованием, но отдельные клетки увеличиваются в размерах, образуют утолщенные оболочки и превращаются в диплоидные хламидоспоры. При прорастании хламидоспоры ее ядро мейотически делится, и непосредственно из хламидоспоры при ее прорастании образуется аск с 1-2 игловидными аскоспорами. Зрелые аскоспоры прорастают и дают начало гаплоидным почкующимся клеткам. Гаплоидные клетки после копуляции восстанавливают диплофазу. Все эти стадии довольно стабильны, поэтому в культурахMetschnikowia pulcherrimaприсутствуют одновременно мелкие гаплоидные клетки, более крупные диплоидные, а такжехламидоспоры.
Базидиомицетовые дрожжи
В отличие от аскоспоровых дрожжей, все дрожжи, которые относят к базидиомицетам, образуют в ходе полового размножения специальные клетки - базидии, несущие экзогенные споры (базидиоспоры). Плазмогамия и кариогамия, как правило, разделены во времени и значительную часть жизненного цикла занимает дикариотическая фаза, представленнаядикариотическим мицелием с пряжками. Большинство видов гетероталличны и поэтому многие из них долгое время были известны только в гаплофазе как несовершенные дрожжи.Жизненные циклы базидиомицетовых дрожжейтакже достаточно разнообразны.
Базидиомицетовые грибы, которые имеют дрожжевую стадию в цикле развития, часто называют гетеробазидиомицетами. Это название было предложено Н.Патулляром еще в 1900 г. Основной признак таких грибов - наличие септированной или глубокорасчлененной базидии. Сейчас этот термин сохраняют для диморфныхбазидиомицетовых грибов с гаплоидной дрожжевой стадией. Ихдикариотический мицелийчасто паразитирует на растениях или плодовых телах других грибов.
Активное исследование жизненных циклов базидиомицетовых дрожжей началось лишь в конце 60-х - начале 70-х годов XX в., после того как японский миколог Исао Банно опубликовал результаты открытия полового циклау хорошо известных и ранее считавшихся несовершенными красных дрожжейRhodotorula glutinis. Ему удалось подобратьтипы спариваниясреди большого числа проверенных штаммов, и в результате их скрещивания он наблюдал образование дикариотического мицелия, на котором формировались структуры, морфологически сходные схламидоспорами, то есть крупные клетки с сильно утолщенными оболочками и большим запасом внутриклеточных липидов. Именно в них затем происходило слияние двух ядер и последующиймейозс восстановлением гаплоидного состояния. Эти клетки получили названиетелиоспор. При прорастании они образовывали промицелий с поперечными септами, делящими его на 4 клетки. Эти клетки отпочковывали гаплоидные споридии. Всю структуру можно было отождествить сфрагмобазидиямителиоспоровых базидиомицетовых грибов. Весь наблюдаемый цикл оказался очень сходным с циклом развитияголовневых грибов. Совершенная стадияRhodotorula glutinisбыла названа БанноRhodosporidium toruloides. Позже сходный цикл был описан для некоторыхбаллистоспоровыхдрожжей и для нескольких видов несовершенного родаCandida. Их телиоспоровые стадии получили соответственно родовые названияSporidiobolusиLeucosporidium. Сейчас похожие полные или не завершенные жизненные циклы известны для многих базидиомицетовых дрожжей. Они различаются формойтелиоспор, характером их прорастания иморфологией базидий.
Другой тип жизненного циклабазидиомицетов с дрожжевой анаморфой известен для дрожалковых грибов порядкаTremellales. Эти грибы известны достаточно давно, так как в онтогенезе этих организмов есть стадия образования макроскопических плодовых тел, которые в природе обычно развиваются на старой древесине, на стволах мертвых деревьев. То, что базидиоспоры дрожалковых грибов способны к почкованию, было замечено микологами еще в конце XIX в., однако наличие самостоятельной дрожжевой фазы в цикле развития тремелловых было установлено после детального изучения видов родаTremellaв 1960-х гг. В культурах отдельные базидиоспоры гетероталличных видов дают начало дрожжевым стадиям. Мицелий обычно развивается только при смешивании штаммов совместимых типов спаривания. Смешение соответствующих типов спаривания приводит к быстрой реакции клеток на выделяющиеся феромоны; почкование прекращается, развиваются конъюгационные трубки, через которые клетки копулируют, происходит плазмогамия и развивается мицелиальнаядикариотическая стадия. У многих видов в подходящих условиях вскоре после формирования дикариотического мицелия начинается развитие плодовых тел.Плодовые телаобычно студенистые или хрящеватые, в виде бесформенных комочков или корочек. В плодовых телах образуются 2-4-клеточныегетеробазидии. Освободившиеся базидиоспоры могут размножаться вегетативно почкованием или формированиембаллистоспор.
В 70-х годах американской исследовательницей Квон-Чанг был обнаружен жизненный циклс образованием несептированных и нерасчлененных базидий у дрожжейCryptococcus neoformans, которые долгое время были известны только в несовершенной форме и привлекали большое внимание медицинских микологов как возбудители опасной болезни -криптококкоза. Серотипы этого вида, обозначаемые A, B, C и D, оказались штаммами разных типов спаривания, которые при скрещивании формировалидикариотический мицелийс несептированными базидиями -голобазидиями. На базидии возникали апикальные вздутия и на них формировались гаплоидные базидиоспоры путем базипетального почкования с образованием цепочек. Совершенная стадия этих дрожжей была названаFilobasidiella neoformans. Позже подобные жизненные циклы были найдены и у некоторых других представителей родаCryptococcus, в частности уCryptococcus uniguttulatus(=Filobasidium uniguttulatum).
Характеристики жизненного цикла играют большую роль в систематике базидиомицетовых дрожжей. Однако очень часто у базидиомицетовых дрожжей, изолируемых из природных местообитаний, не удается наблюдать полный жизненный цикл при культивировании на лабораторных средах, поэтому их идентификация и классификация вызывают большие трудности. Значительные успехи в систематике таких анаморфных базидиомицетовых дрожжей достигнуты в последние десятилетия, особенно в связи с внедрением в таксономическую практику молекулярно-биологических методов. Как оказалось, группирование базидиомицетовых дрожжей на основе молекулярно-биологических признаков, в частности нуклеотидных последовательностей рРНК, часто не совпадает с характеристиками их жизненного цикла, особенно такими, как способность к образованию телиоспор, морфология базидий. Поэтому классификация базидиомицетовых дрожжей еще далека от завершенности и требует дальнейших исследований.
studfiles.net
Дрожжи - Аскомицетные и базидиомицетные дрожжи
Химия - Дрожжи - Аскомицетные и базидиомицетные дрожжи
01 марта 2011Оглавление:1. Дрожжи2. История3. Аскомицетные и базидиомицетные дрожжи4. Распространение5. Типы спаривания6. Применение7. Дрожжи как фактор порчи пищевых продуктов8. Патогенные дрожжи
Различить дрожжи, принадлежащие к разным отделам грибов можно как по характеристикам их жизненного цикла, так и без его наблюдения по признакам аффинитета. К ним относится: синтез каротиноидов, тип убихинонов, тип почкования, содержание ГЦ пар в ДНК, наличие уреазы и др.
Типичное разделение
Аскомицеты
- Saccharomycotina
- Taphrinomycotina
Базидиомицеты
Особенности метаболизма
Дрожжи являются хемоорганогетеротрофами и используют органические соединения как для получения энергии, так и в качестве источника углерода. Им необходим кислород для дыхания, однако при его отсутствии многие виды способны получать энергию за счёт брожения с выделением спиртов. В отличие от бактерий, среди дрожжей нет облигатных анаэробов, гибнущих при наличии кислорода в среде. При пропускании воздуха через сбраживаемый субстрат дрожжи прекращают брожение и начинают дышать, потребляя кислород и выделяя углекислый газ. Это ускоряет рост дрожжевых клеток. Однако даже при доступе кислорода в случае высокого содержания глюкозы в среде дрожжи начинают её сбраживать.
Дрожжи достаточно требовательны к условиям питания. В анаэробных условиях дрожжи могут использовать в качестве источника энергии только углеводы, причём в основном гексозы и построенные из них олигосахариды. Некоторые виды усваивают и пентозы, например, ксилозу. Schwanniomyces occidentalis и Saccharomycopsis fibuliger способны сбраживать крахмал, Kluyveromyces fragilis инулин. В аэробных условиях круг усваиваемых субстратов шире: помимо углеводов в него входят также жиры, углеводороды, ароматические и одноуглеродные соединения, спирты, органические кислоты. Гораздо больше видов способно использовать пентозы в аэробных условиях. Тем не менее, сложные соединения для большинства дрожжей недоступны.
Источниками азота для всех дрожжей могут быть соли аммония, примерно половина видов имеет нитратредуктазу и может усваивать нитраты. Пути усвоения мочевины различны у аскомицетовых и базидиомицетовых дрожжей. Аскомицетовые сначала карбоксилируют её, затем гидролизуют, базидиомицетовые сразу гидролизуют уреазой.
Для практического применения важны продукты вторичного метаболизма дрожжей, выделяемые в малых количествах в среду: сивушные масла, ацетоин, диацетил, масляный альдегид, изоамиловый спирт, диметилсульфид и др. Именно от них зависят органолептические свойства полученных с помощью дрожжей продуктов.
Просмотров: 5864
4108.ru
Дрожжи - Аскомицетные и базидиомицетные дрожжи
Биология - Дрожжи - Аскомицетные и базидиомицетные дрожжи
09 февраля 2011Оглавление:1. Дрожжи2. История3. Аскомицетные и базидиомицетные дрожжи4. Распространение5. Типы спаривания6. Применение7. Дрожжи как фактор порчи пищевых продуктов8. Патогенные дрожжи
Различить дрожжи, принадлежащие к разным отделам грибов можно как по характеристикам их жизненного цикла, так и без его наблюдения по признакам аффинитета. К ним относится: синтез каротиноидов, тип убихинонов, тип почкования, содержание ГЦ пар в ДНК, наличие уреазы и др.
Типичное разделение
Аскомицеты
- Saccharomycotina
- Taphrinomycotina
Базидиомицеты
Особенности метаболизма
Дрожжи являются хемоорганогетеротрофами и используют органические соединения как для получения энергии, так и в качестве источника углерода. Им необходим кислород для дыхания, однако при его отсутствии многие виды способны получать энергию за счёт брожения с выделением спиртов. В отличие от бактерий, среди дрожжей нет облигатных анаэробов, гибнущих при наличии кислорода в среде. При пропускании воздуха через сбраживаемый субстрат дрожжи прекращают брожение и начинают дышать, потребляя кислород и выделяя углекислый газ. Это ускоряет рост дрожжевых клеток. Однако даже при доступе кислорода в случае высокого содержания глюкозы в среде дрожжи начинают её сбраживать.
Дрожжи достаточно требовательны к условиям питания. В анаэробных условиях дрожжи могут использовать в качестве источника энергии только углеводы, причём в основном гексозы и построенные из них олигосахариды. Некоторые виды усваивают и пентозы, например, ксилозу. Schwanniomyces occidentalis и Saccharomycopsis fibuliger способны сбраживать крахмал, Kluyveromyces fragilis инулин. В аэробных условиях круг усваиваемых субстратов шире: помимо углеводов в него входят также жиры, углеводороды, ароматические и одноуглеродные соединения, спирты, органические кислоты. Гораздо больше видов способно использовать пентозы в аэробных условиях. Тем не менее, сложные соединения для большинства дрожжей недоступны.
Источниками азота для всех дрожжей могут быть соли аммония, примерно половина видов имеет нитратредуктазу и может усваивать нитраты. Пути усвоения мочевины различны у аскомицетовых и базидиомицетовых дрожжей. Аскомицетовые сначала карбоксилируют её, затем гидролизуют, базидиомицетовые сразу гидролизуют уреазой.
Для практического применения важны продукты вторичного метаболизма дрожжей, выделяемые в малых количествах в среду: сивушные масла, ацетоин, диацетил, масляный альдегид, изоамиловый спирт, диметилсульфид и др. Именно от них зависят органолептические свойства полученных с помощью дрожжей продуктов.
Просмотров: 27610
| Дейтеромицеты |
www.muldyr.ru
Базидиомицеты - Грибы - Общая микробиология - Статьи
Базидиомицеты считаются наиболее высокоразвитой группой грибов. Характерный для них орган спороношения – базидия - по своему расположению (это клетка на конце гифы) соответствует аску. От базидии отшнуровываются, как правило, четыре одноядерные гаплоидные базидиоспоры. Подобно аскоспорам, они образуются в результате плазмогамии, кариогамии и мейоза. Два последних процесса протекают соответственно в аске или в базидии.
Мицелий состоит из гиф с поперечными перегородками. Белые нити, которые заметны невооруженному глазу в лесной подстилке, - это пучки гиф, окруженные «корой»; их называют ризоморфами.
Цикл развития гименомицетов. Из проросшей базидиоспоры образуется первичный мицелий, состоящий из гиф, разделенных перегородками на одноядерные клетки. Вторичный, дикариофитный, мицелий возникает в тех случаях, когда встречаются гифы двух совместимых штаммов и их одноядерные протопласты сливаются (плазмогамия). При каждом клеточном делении происходит сопряженное деление обоих ядер. У многих базидиомицетов деление клеток и ядер сопровождается образованием так называемой пряжки (рис. 5.14). Такой механизм обеспечивает получение новой клеткой по одному дочернему ядру каждого типа. У клетки, готовой к делению, между ядрами а и b образуется нечто вроде крючка, который загибается назад; переднее ядро b переходит в этот «крючок» (пряжку), после чего оба ядра делятся. За тем передняя часть клетки, содержащая дочерние ядра а и b, отделяется от задней части поперечной перегородкой. Одновременно пряжка сливается с исходной клеткой, в которую при этом возвращается ядро b. Передняя клетка отделена перегородкой также и от пряжки. В результате обе клетки получают по одному ядру типа а и одному ядру типа b.
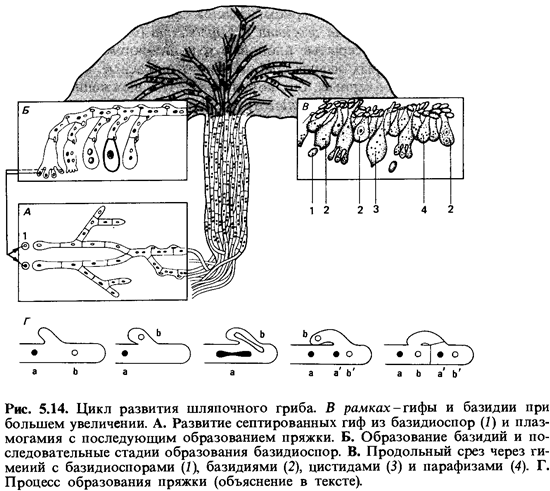
На дикариофитном мицелии вырастают в типичных случаях сложные плодовые тела. Это хорошо известные шляпочные грибы, трутовики и т.п. Образование базидии происходит в специальном слое такого плодового тела - в так называемом гимении, который состоит из базидии и стерильных гиф (парафиз и цистид). Базидия образуется в результате увеличения концевой клетки; два ядра этой клетки сливаются (кариогамия). Затем ядро зиготы претерпевает редукционное деление с образованием четырех гаплоидных ядер. Одновременно формируются четыре стеригмы с зачатками базидиоспор, в которые проникают ядра.
Базидиоспоры активно выбрасываются плодовым телом. При массовом скоплении базидиоспор можно заметить, что они пигментированы. Их окраска - коричневая, фиолетовая, черная, цвета охры и т. д. - служит важным диагностическим признаком.
Описанный выше цикл развития (с некоторыми модификациями) типичен для гомобазидиомицетов. Для них характерно наличие именно таких базидий. По расположению гимения различают гименомицеты и гастеромицеты. К гименомицетам относятся шляпочные грибы (пластинчатые, трубчатые, игольчатые и т.д.), трутовики и коралловидные формы. У гастеромицетов (дождевики, Geaster, Phyllaceae и др.) плодовое тело не открывается и гимений не обнажается. Базидиоспоры освобождаются только после распада гимения.
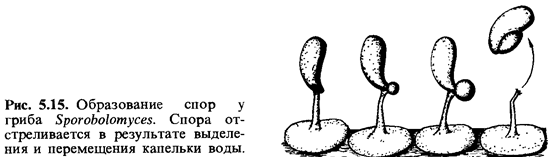
К гетеробазидиомицетам относятся дрожалковые (Tremellales), ржавчинные (Uredinales) и головневые (Ustilaginales) грибы. По своему строению их базидии отличаются от базидий гомобазидиомицетов. У некоторых групп пробазидия окружена твердой стенкой и ведет себя как покоящаяся спора. К сожалению, мы не можем здесь углубиться в такие интересные подробности, как различные модификации базидий, приспособление к условиям среды, к вегетативному циклу растений-хозяев и т.п. Упомянем, однако, еще об одном небольшом семействе грибов, сходном с дрожжами, - о Sporobolomycetaceae. Эти грибы образуют активно отбрасываемые конидии, сидящие на стеригмах; такие конидии называют баллистоспорами. Sporobolomycessalmonicolor, подобно дрожжам, размножается почкованием. Каждая его клетка может образовать стеригму, на которой появляется спора, по форме напоминающая почку. При созревании у нижнего конца споры, в месте ее прикрепления, выделяется капелька воды, и спора подбрасывается на высоту около 0,1 мм (рис. 5.15). Видимо, при помощи такого механизма происходит выбрасывание базидиоспор у большинства форм со свободно сидящими базидиями (гименомицетов).
micro.moy.su

















 |  |  |  |  |  |  |  |  |  |  |  |  |  |  |
 Пример видео 3 Пример видео 3 |  Пример видео 2 Пример видео 2 |  Пример видео 6 Пример видео 6 |  Пример видео 1 Пример видео 1 |  Пример видео 5 Пример видео 5 |  Пример видео 4 Пример видео 4 |
Администрация муниципального образования «Городское поселение – г.Осташков»